

doi: 10.1128/JB.186.12.4014-4018.2004.
Cross-regulation in Vibrio parahaemolyticus: compensatory activation of polar flagellar genes by the lateral flagellar regulator LafK
Affiliations
- PMID: 15175315
- PMCID: PMC419961
- DOI: 10.1128/JB.186.12.4014-4018.2004
Item in Clipboard
Cross-regulation in Vibrio parahaemolyticus: compensatory activation of polar flagellar genes by the lateral flagellar regulator LafK
J Bacteriol.
2004 Jun.
Abstract
Gene organization and hierarchical regulation of the polar flagellar genes of Vibrio parahaemolyticus, Vibrio cholerae, and Pseudomonas aeruginosa appear highly similar, with one puzzling difference. Two sigma(54)-dependent regulators are required to direct different classes of intermediate flagellar gene expression in V. cholerae and P. aeruginosa, whereas the V. parahaemolyticus homolog of one of these regulators, FlaK, appears dispensable. Here we demonstrate that there is compensatory activation of polar flagellar genes by the lateral flagellar regulator LafK.
Figures
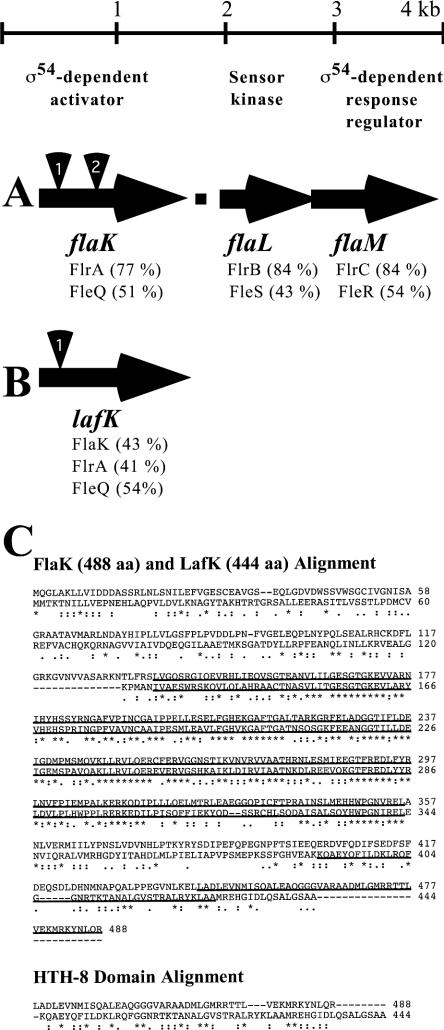
Polar and lateral flagellar regulators. (A) Physical map of the locus encoding the V. parahaemolyticus polar flagellar regulators. A potential rho-independent transcriptional terminator (closed square) occurs between the coding regions for flaK and flaL. Homologous gene clusters are found in other bacteria such as V. cholerae (flrA flrBC) and P. aeruginosa (fleQ fleSR). The percent identities obtained in a pairwise BLAST analysis (22) with the V. parahaemolyticus homolog are provided in parentheses. FlaK, FlaM, and their homologs appear to be transcription factors with potential σ54-interacting domains. FlaL and its homologs are potential sensor kinases. FlaM and its homologs, in addition to containing σ54-interacting domains, are potential response regulators. The positions of insertions introduced into the flaK coding region are indicated: a chloramphenicol resistance cassette was inserted into the NsiI site at position bp 200 (to make flaK1) and the HpaI site at bp 640 (to make flaK2). (B) The lateral flagellar regulatorygene lafK also encodes a potential σ54-interacting transcription factor. LafK is essential for swarming but is not required for swimming motility. The position of the insertion used to disrupt lafK is indicated: a gentamicin resistance lacZ cassette was introduced into the SacI site at bp 154 of the coding region. The percent identities obtained in a BLAST analysis of LafK with its polar flagellar homolog from each organism are shown. (C) Clustal W alignments of FlaK and LafK (http://www.ebi.ac.uk/clustalw ). The alignment symbols denoting degree of conservation are as follows: *, residues are identical; :, conserved substitutions; and ., semiconserved substitutions. The top alignment compares FlaK (488 amino acids) in the top line and LafK (444 amino acids) in the lower line. The centrally located, conserved σ54-interacting domains (pfam00158) are underlined for both regulatory proteins. Also underlined are N terminally located conserved HTH_8 domains (pfam02954) common to bacterial regulatory proteins in the FIS family. The bottom alignment shows the comparison of the HTH_8 domains.
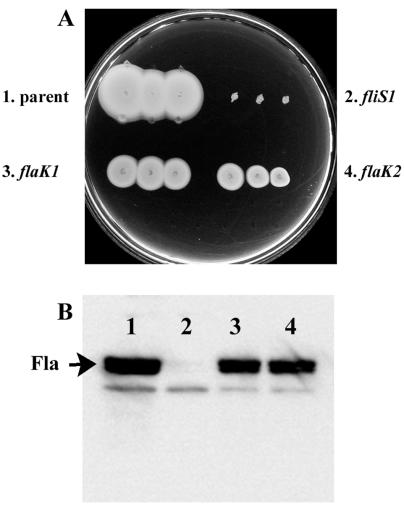
Introduction of mutations in flaK does not abolish swimming motility. (A) Swimming motility in semisolid tryptone motility agar (1% tryptone, 2% NaCl, 0.32% agar) after overnight incubation at room temperature. Three single colonies of each strain were inoculated. (B) Immunoblot of polar flagellin (Fla) profiles with samples prepared from strains grown in heart infusion broth. Immunoblotting conditions have been described (3) except that blots were incubated with primary antiserum for 14 h. The longer incubation period allowed detection of a cross-reacting cellular protein that serves as a loading control. All strains were derived from strain LM1017, which contains a defect in the lateral flagellar hook gene; therefore there is no contribution to motility from the lateral flagella (Fla+ Laf−). Strains: lane 1, LM1017 (flgE313L); lane 2, LM4348 (fliS1::Camr in LM1017); lane 3, LM4347 (flaK1::Camr in LM1017); and lane 4, LM4349 (flaK2::Camr in LM1017). The nonmotile control strain LM4348 contains a defect in the polar flagellar chaperone fliS (formerly named flaJ) (21).
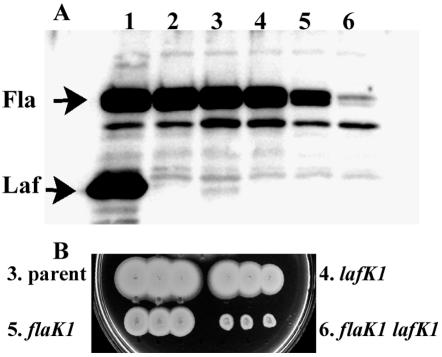
flaK or lafK mutants swim, but flaK lafK mutants are impaired for swimming motility. (A) Polar (Fla) and lateral (Laf) flagellin profiles in immunoblots with samples prepared from strains grown overnight on HI plates. Strains: lane 1, LM5674 (wild type, Fla+ Laf+); lane 2, LM7011 (lafK1::Genr in LM5674); lane 3, LM1017 (flgE313L); lane 4, LM7010 (lafK1::Genr in LM1017); lane 5, LM4347 (flaK1::Camr in LM1017); and lane 6, LM6856 (lafK1::Genr flaK1::Camr in LM1017). (B) Swimming motility in semisolid tryptone motility agar. Plates were inoculated with three single colonies of each strain and incubated at room temperature overnight. Strains are numbered as for panel A.
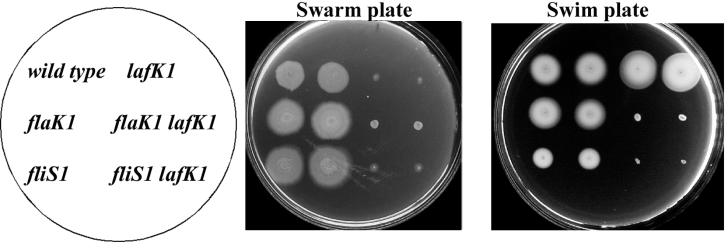
In the wild-type background, the flaK mutation has a phenotype only in the context of a lafK mutation. Two single colonies of each strain were inoculated on a swarm plate (2.5% heart infusion, 2% NaCl, 1.5% agar [DIFCO]) and into a swim plate (semisolid tryptone motility agar; 0.325% agar [Difco]). Plates were incubated at room temperature overnight. Strains: LM5674 (wild type), LM7011 (lafK1::Genr), LM4471 (flaK1::Camr), LM7305 (flaK1::Camr lafK1::Genr), LM4472 (fliS1::Camr), and LM7395 (fliS1::Camr lafK1::Genr).
Similar articles
-
Lateral flagellar gene system of Vibrio parahaemolyticus.J Bacteriol. 2003 Aug;185(15):4508-18. doi: 10.1128/JB.185.15.4508-4518.2003. J Bacteriol. 2003. PMID: 12867460 Free PMC article.
-
Deciphering bacterial flagellar gene regulatory networks in the genomic era.Adv Appl Microbiol. 2009;67:257-95. doi: 10.1016/S0065-2164(08)01008-3. Adv Appl Microbiol. 2009. PMID: 19245942 Review.
-
Dual flagellar systems enable motility under different circumstances.J Mol Microbiol Biotechnol. 2004;7(1-2):18-29. doi: 10.1159/000077866. J Mol Microbiol Biotechnol. 2004. PMID: 15170400 Review.
-
Vibrio parahaemolyticus FlaJ, a homologue of FliS, is required for production of a flagellin.Mol Microbiol. 1996 Apr;20(1):137-49. doi: 10.1111/j.1365-2958.1996.tb02496.x. Mol Microbiol. 1996. PMID: 8861212
-
The novel sigma54- and sigma28-dependent flagellar gene transcription hierarchy of Vibrio cholerae.Mol Microbiol. 2001 Mar;39(6):1595-609. doi: 10.1046/j.1365-2958.2001.02348.x. Mol Microbiol. 2001. PMID: 11260476
Cited by
-
Transcriptome Analysis Reveals Cross-Talk between the Flagellar Transcriptional Hierarchy and Secretion System in Plesiomonas shigelloides.Int J Mol Sci. 2024 Jul 5;25(13):7375. doi: 10.3390/ijms25137375. Int J Mol Sci. 2024. PMID: 39000482 Free PMC article.
-
Environment-directed activation of the Escherichia coliflhDC operon by transposons.Microbiology (Reading). 2017 Apr;163(4):554-569. doi: 10.1099/mic.0.000426. Epub 2017 Apr 12. Microbiology (Reading). 2017. PMID: 28100305 Free PMC article.
-
Transcriptional hierarchy of Aeromonas hydrophila polar-flagellum genes.J Bacteriol. 2011 Oct;193(19):5179-90. doi: 10.1128/JB.05355-11. Epub 2011 Jul 22. J Bacteriol. 2011. PMID: 21784933 Free PMC article.
-
Characterization of the relationship between polar and lateral flagellar structural genes in the deep-sea bacterium Shewanella piezotolerans WP3.Sci Rep. 2016 Dec 22;6:39758. doi: 10.1038/srep39758. Sci Rep. 2016. PMID: 28004809 Free PMC article.
-
Characterizing the Adherence Profiles of Virulent Vibrio parahaemolyticus Isolates.Microb Ecol. 2018 Jan;75(1):152-162. doi: 10.1007/s00248-017-1025-8. Epub 2017 Jul 17. Microb Ecol. 2018. PMID: 28717834
References
-
- Aldridge, P., and K. T. Hughes. 2002. Regulation of flagellar assembly. Curr. Opin. Microbiol. 5:160-165. - PubMed
-
- Correa, N. E., C. M. Lauriano, R. McGee, and K. E. Klose. 2000. Phosphorylation of the flagellar regulatory protein FlrC is necessary for Vibrio cholerae motility and enhanced colonization. Mol. Microbiol. 35:743-755. - PubMed
-
- Dasgupta, N., M. C. Wolfgang, A. L. Goodman, S. K. Arora, J. Jyot, S. Lory, and R. Ramphal. 2003. A four-tiered transcriptional regulatory circuit controls flagellar biogenesis in Pseudomonas aeruginosa. Mol. Microbiol. 50:809-824. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
