

The topoisomerase IV ParC subunit colocalizes with the Caulobacter replisome and is required for polar localization of replication origins
- PMID: 15178756
- PMCID: PMC438962
- DOI: 10.1073/pnas.0402567101
The topoisomerase IV ParC subunit colocalizes with the Caulobacter replisome and is required for polar localization of replication origins
Abstract
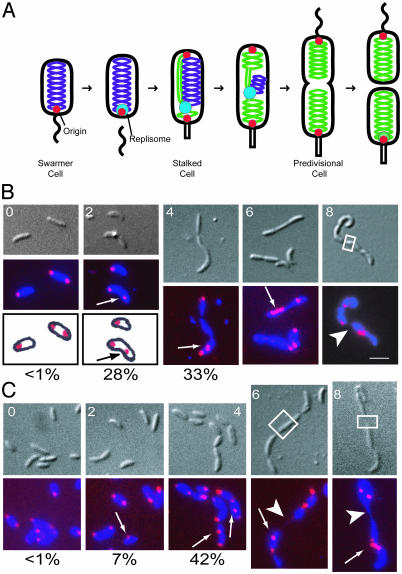
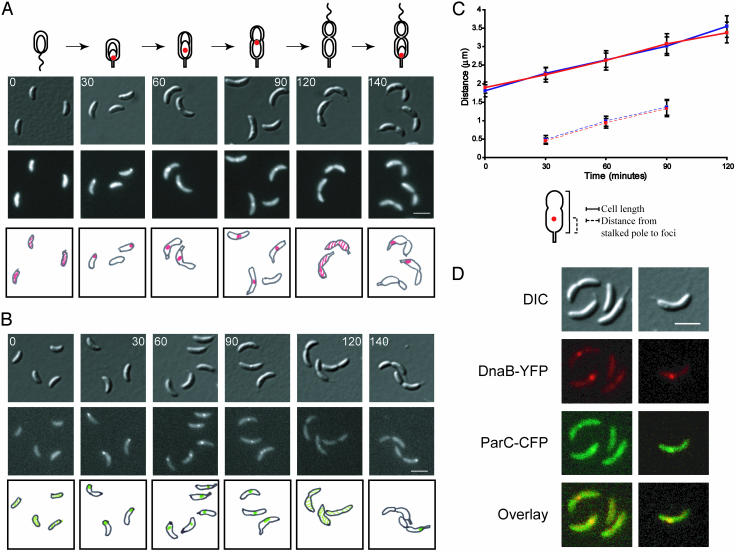
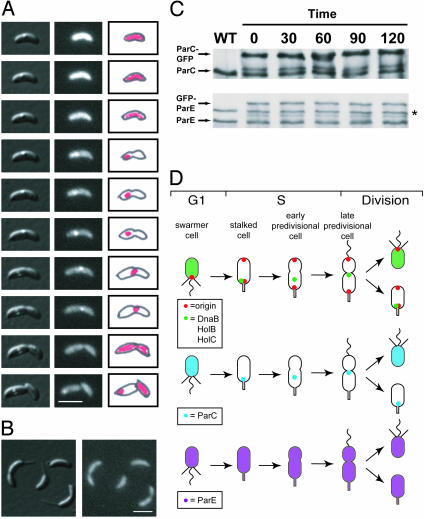
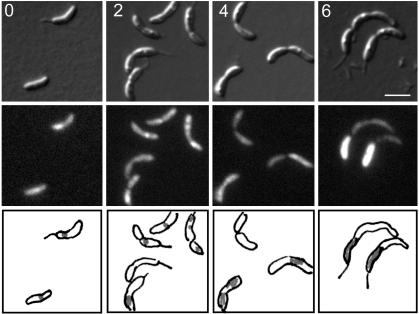
The process of bacterial DNA replication generates chromosomal topological constraints that are further confounded by simultaneous transcription. Topoisomerases play a key role in ensuring orderly replication and partition of DNA in the face of a continuously changing DNA tertiary structure. In addition to topological constraints, the cellular position of the replication origin is strictly controlled during the cell cycle. In Caulobacter crescentus, the origin of DNA replication is located at the cell pole. Upon initiation of DNA replication, one copy of the duplicated origin sequence rapidly appears at the opposite cell pole. To determine whether the maintenance of DNA topology contributes to the dynamic positioning of a specific DNA region within the cell, we examined origin localization in cells that express temperature-sensitive forms of either the ParC or ParE subunit of topoisomerase (Topo) IV. We found that in the absence of active Topo IV, replication initiation can occur but a significant percent of replication origins are either no longer moved to or maintained at the cell poles. During the replication process, the ParC subunit colocalizes with the replisome, whereas the ParE subunit is dispersed throughout the cell. However, an active ParE subunit is required for ParC localization to the replisome as it moves from the cell pole to the division plane during chromosome replication. We propose that the maintenance of DNA topology throughout the cell cycle contributes to the dynamic positioning of the origin sequence within the cell.
Figures
Similar articles
-
Cell cycle expression and transcriptional regulation of DNA topoisomerase IV genes in caulobacter.J Bacteriol. 1999 Jun;181(11):3321-9. doi: 10.1128/JB.181.11.3321-3329.1999. J Bacteriol. 1999. PMID: 10348842 Free PMC article.
-
A moving DNA replication factory in Caulobacter crescentus.EMBO J. 2001 Sep 3;20(17):4952-63. doi: 10.1093/emboj/20.17.4952. EMBO J. 2001. PMID: 11532959 Free PMC article.
-
Requirement of topoisomerase IV parC and parE genes for cell cycle progression and developmental regulation in Caulobacter crescentus.Mol Microbiol. 1997 Dec;26(5):897-910. doi: 10.1046/j.1365-2958.1997.6242005.x. Mol Microbiol. 1997. PMID: 9426128
-
Control of chromosome replication in caulobacter crescentus.Annu Rev Microbiol. 2002;56:625-56. doi: 10.1146/annurev.micro.56.012302.161103. Epub 2002 Jan 30. Annu Rev Microbiol. 2002. PMID: 12142494 Review.
-
Setting the pace: mechanisms tying Caulobacter cell-cycle progression to macroscopic cellular events.Curr Opin Microbiol. 2004 Apr;7(2):192-7. doi: 10.1016/j.mib.2004.02.012. Curr Opin Microbiol. 2004. PMID: 15063858 Review.
Cited by
-
Mutational Diversity in the Quinolone Resistance-Determining Regions of Type-II Topoisomerases of Salmonella Serovars.Antibiotics (Basel). 2021 Nov 26;10(12):1455. doi: 10.3390/antibiotics10121455. Antibiotics (Basel). 2021. PMID: 34943668 Free PMC article. Review.
-
Rapid and sequential movement of individual chromosomal loci to specific subcellular locations during bacterial DNA replication.Proc Natl Acad Sci U S A. 2004 Jun 22;101(25):9257-62. doi: 10.1073/pnas.0402606101. Epub 2004 Jun 3. Proc Natl Acad Sci U S A. 2004. PMID: 15178755 Free PMC article.
-
Sister chromatid interactions in bacteria revealed by a site-specific recombination assay.EMBO J. 2012 Aug 15;31(16):3468-79. doi: 10.1038/emboj.2012.194. Epub 2012 Jul 20. EMBO J. 2012. PMID: 22820946 Free PMC article.
-
Novel Ser79Leu and Ser81Ile substitutions in the quinolone resistance-determining regions of ParC topoisomerase IV and GyrA DNA gyrase subunits from recent fluoroquinolone-resistant Streptococcus pneumoniae clinical isolates.Antimicrob Agents Chemother. 2005 Jun;49(6):2479-86. doi: 10.1128/AAC.49.6.2479-2486.2005. Antimicrob Agents Chemother. 2005. PMID: 15917550 Free PMC article.
-
The SMC complex MukBEF recruits topoisomerase IV to the origin of replication region in live Escherichia coli.mBio. 2014 Feb 11;5(1):e01001-13. doi: 10.1128/mBio.01001-13. mBio. 2014. PMID: 24520061 Free PMC article.
References
-
- Teleman, A. A., Graumann, P. L., Lin, D. C., Grossman, A. D. & Losick, R. (1998) Curr. Biol. 8, 1102-1109. - PubMed
-
- Lewis, P. J. & Errington, J. (1997) Mol. Microbiol. 25, 945-954. - PubMed
-
- Webb, C. D., Teleman, A., Gordon, S., Straight, A., Belmont, A., Lin, D. C., Grossman, A. D., Wright, A. & Losick, R. (1997) Cell 88, 667-674. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
