

Cytokine stimulation of aerobic glycolysis in hematopoietic cells exceeds proliferative demand
- PMID: 15180958
- PMCID: PMC4458073
- DOI: 10.1096/fj.03-1001fje
Cytokine stimulation of aerobic glycolysis in hematopoietic cells exceeds proliferative demand
Abstract
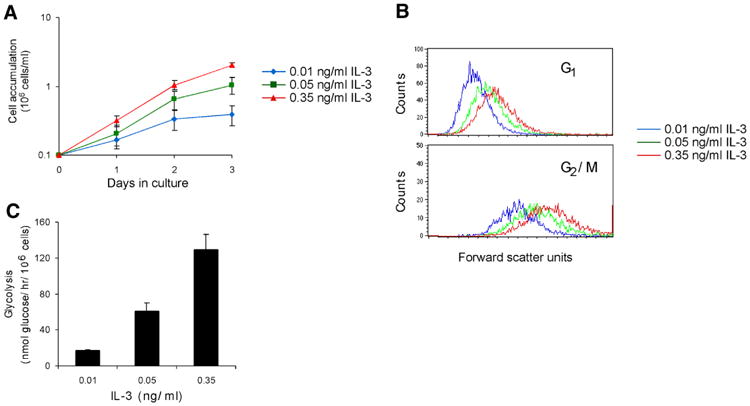
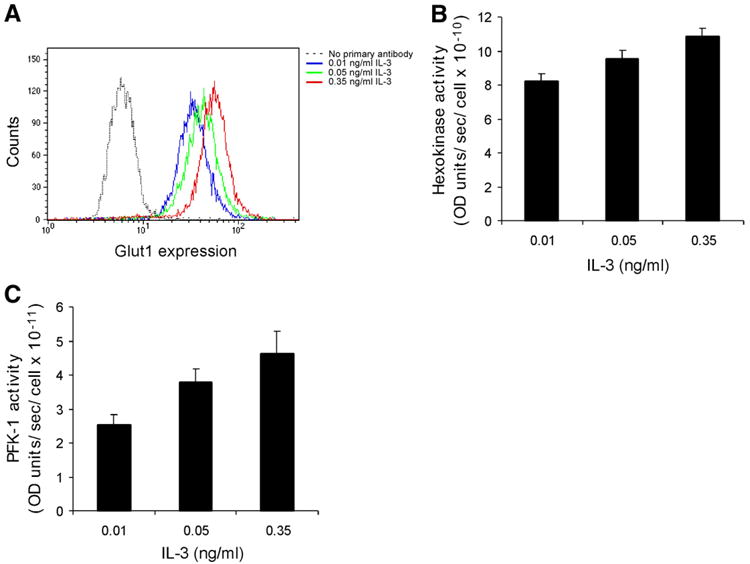
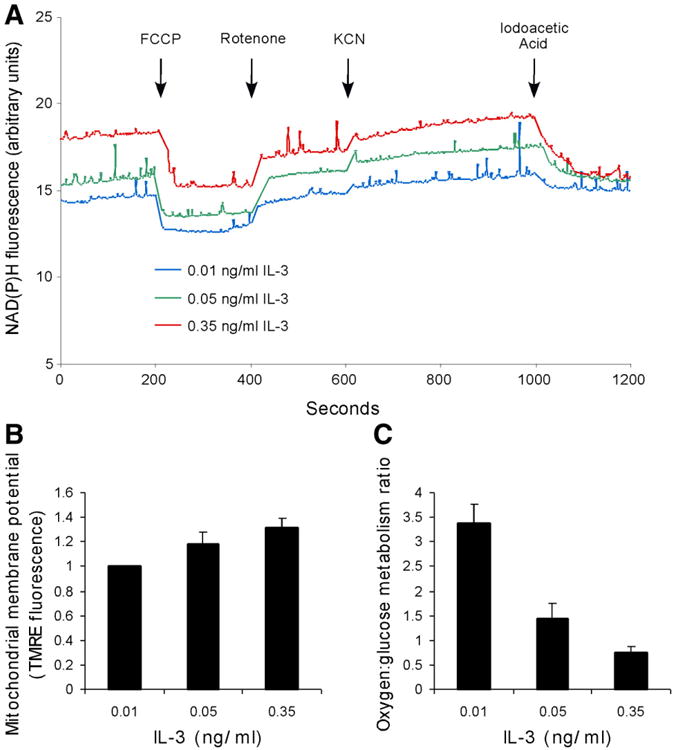
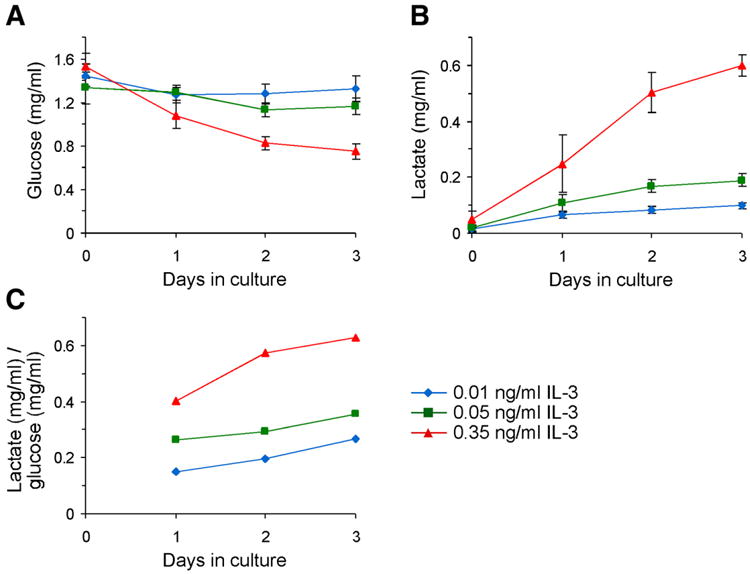
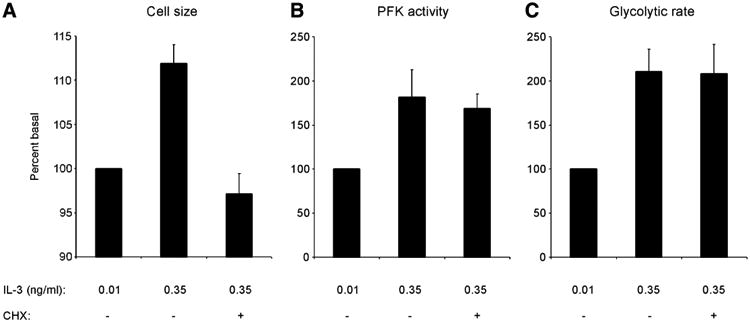
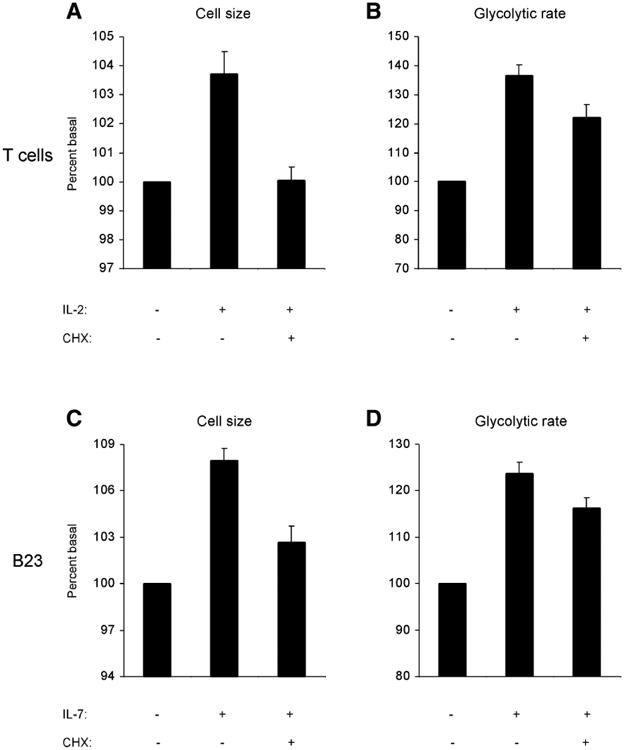
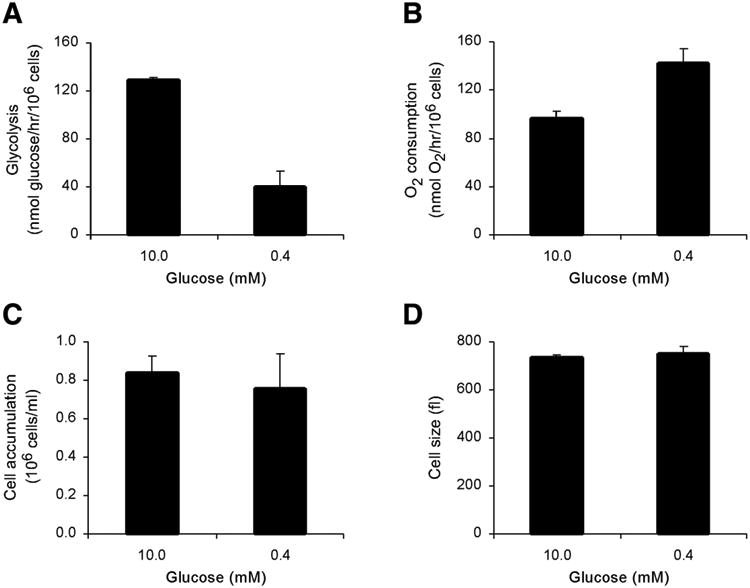
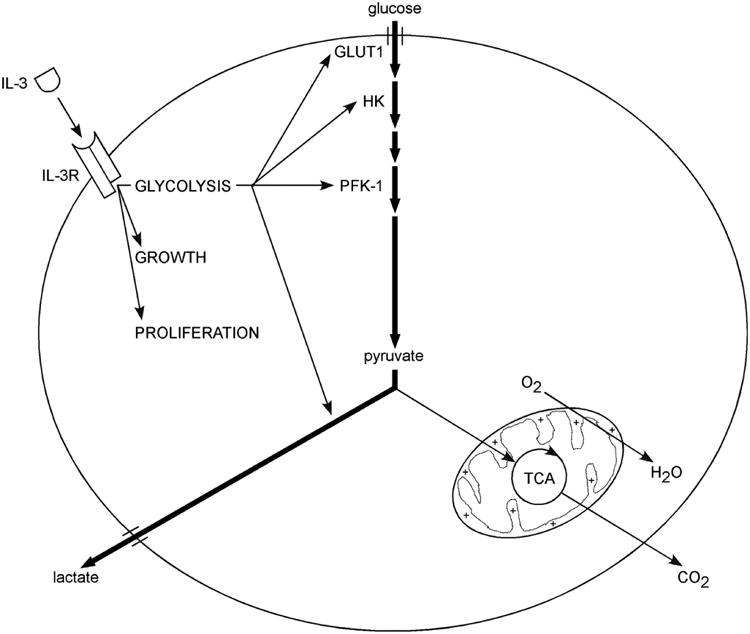
The relationship between growth factor-dependent cell growth and proliferation and the up-regulation of cellular metabolism required to support these processes remains poorly defined. Here, we demonstrate that cell growth, proliferation, and glucose metabolism are coordinately regulated by interleukin-3 (IL-3) in cytokine-dependent cells. Surprisingly, glycolytic activity is stimulated to a greater extent than would be expected based on the rate of cell growth or proliferation. IL-3 signaling exerts a direct effect on glycolytic commitment independent of cell growth control. These results are not restricted to IL-3 as the cytokines IL-7 and IL-2 have similar effects on glucose metabolism when assayed in factor-dependent cell lines or primary lymphocytes, respectively. Growth factor stimulation leads cells to consume less oxygen and produce more lactate per glucose, indicative of conversion from oxidative to glycolytic metabolism. The enforced rate of glucose metabolism is in excess of that required to support cell growth; accordingly, if extracellular glucose is reduced, cells retain the ability to grow and proliferate by derepressing oxidative metabolism. These data suggest that the high rate of glycolysis observed in response to growth factor stimulation is a primary effect rather than a homeostatic response to increased cell growth.
Figures
References
-
- Conlon I, Raff M. Size control in animal development. Cell. 1999;96:235–244. - PubMed
-
- Cooper EH, Barkhan P, Hale AJ. Observations on the proliferation of human leucocytes cultured with phytohaemagglutinin. Br J Haematol. 1963;9:101–111. - PubMed
-
- Sagone AL, Jr, LoBuglio AF, Balcerzak SP. Alterations in hexose monophosphate shunt during lymphoblastic transformation. Cell Immunol. 1974;14:443–452. - PubMed
-
- Roos D, Loos JA. Changes in the carbohydrate metabolism of mitogenically stimulated human peripheral lymphocytes. I Stimulation by phytohaemagglutinin. Biochim Biophys Acta. 1970;222:565–582. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
