

Cell cycle progression after cleavage failure: mammalian somatic cells do not possess a "tetraploidy checkpoint"
- PMID: 15184397
- PMCID: PMC2172377
- DOI: 10.1083/jcb.200403014
Cell cycle progression after cleavage failure: mammalian somatic cells do not possess a "tetraploidy checkpoint"
Abstract
Failure of cells to cleave at the end of mitosis is dangerous to the organism because it immediately produces tetraploidy and centrosome amplification, which is thought to produce genetic imbalances. Using normal human and rat cells, we reexamined the basis for the attractive and increasingly accepted proposal that normal mammalian cells have a "tetraploidy checkpoint" that arrests binucleate cells in G1, thereby preventing their propagation. Using 10 microM cytochalasin to block cleavage, we confirm that most binucleate cells arrest in G1. However, when we use lower concentrations of cytochalasin, we find that binucleate cells undergo DNA synthesis and later proceed through mitosis in >80% of the cases for the hTERT-RPE1 human cell line, primary human fibroblasts, and the REF52 cell line. These observations provide a functional demonstration that the tetraploidy checkpoint does not exist in normal mammalian somatic cells.
Copyright the Rockefeller University Press
Figures
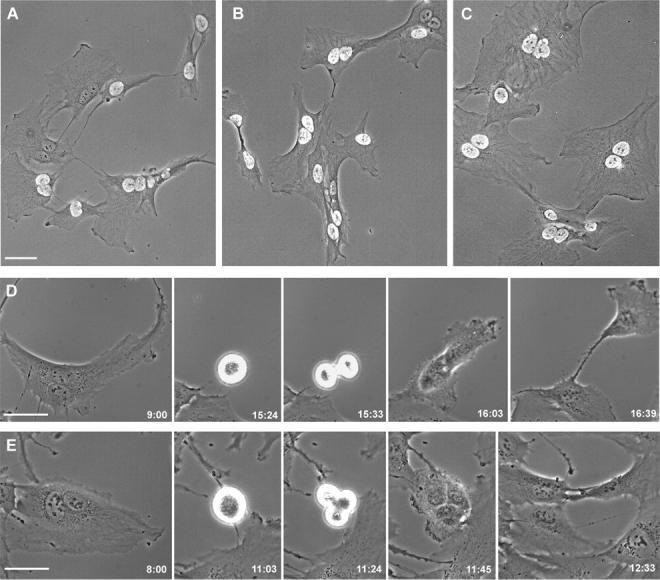
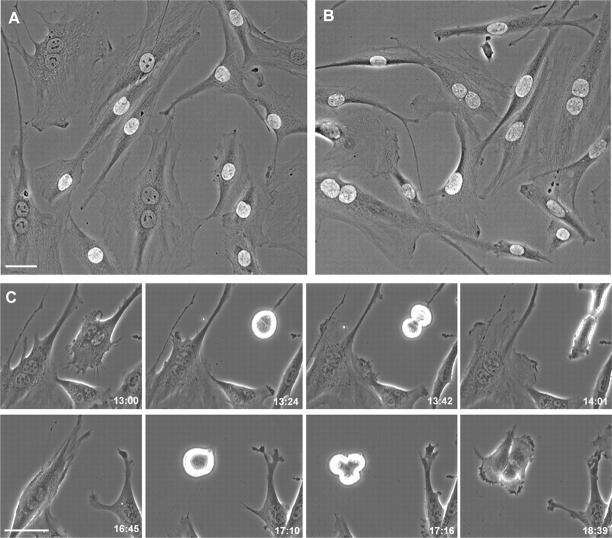

Comment in
-
Triggering p53 after cytokinesis failure.J Cell Biol. 2004 Jun 7;165(5):607-8. doi: 10.1083/jcb.200405089. J Cell Biol. 2004. PMID: 15184396 Free PMC article. Review.
References
-
- Ailenberg, M., and M. Silverman. 2003. Cytochalasin D disruption of actin filaments in 3T3 cells produces an anti-apoptotic response by activating gelatinase A extracellularly and initiating intracellular survival signals. Biochim. Biophys. Acta. 1593:249–258. - PubMed
-
- Brinkley, B.R. 2001. Managing the centrosome numbers game: from chaos to stability in cancer cell division. Trends Cell Biol. 11:18–21. - PubMed
