

Tumor rejection induced by CD70-mediated quantitative and qualitative effects on effector CD8+ T cell formation
- PMID: 15184507
- PMCID: PMC2211777
- DOI: 10.1084/jem.20031111
Tumor rejection induced by CD70-mediated quantitative and qualitative effects on effector CD8+ T cell formation
Abstract
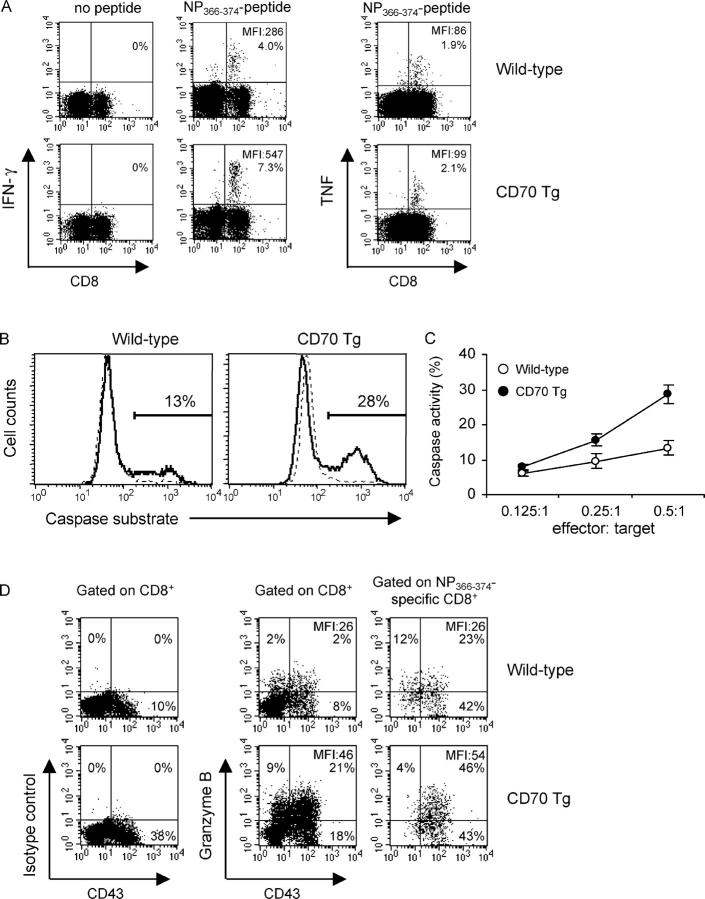
In vivo priming of antigen-specific CD8+ T cells results in their expansion and differentiation into effector T cells followed by contraction into a memory T cell population that can be maintained for life. Recent evidence suggests that after initial antigenic stimulation, the magnitude and kinetics of the CD8+ T cell response are programmed. However, it is unclear to what extent CD8+ T cell instruction in vivo is modulated by costimulatory signals. Here, we demonstrate that constitutive ligation of the tumor necrosis factor receptor family member CD27 by its ligand CD70 quantitatively augments CD8+ T cell responses to influenza virus infection and EL-4 tumor challenge in vivo by incrementing initial expansion and maintaining higher numbers of antigen-specific T cells in the memory phase. Concomitantly, the quality of antigen-specific T cells improved as evidenced by increased interferon (IFN)-gamma production and a greater cytotoxic potential on a per cell basis. As an apparent consequence, the superior effector T cell formation induced by CD70 protected against a lethal dose of poorly immunogenic EL4 tumor cells in a CD8+ T cell- and IFN-gamma-dependent manner. Thus, CD70 costimulation enhances both the expansion and per cell activity of antigen-specific CD8+ T cells.
Figures





References
-
- Sprent, J., and C.D. Surh. 2002. T cell memory. Annu. Rev. Immunol. 20:551–579. - PubMed
-
- Wong, P., and E.G. Pamer. 2003. CD8 T cell responses to infectious pathogens. Annu. Rev. Immunol. 21:29–70. - PubMed
-
- Ahmed, R., and D. Gray. 1996. Immunological memory and protective immunity: understanding their relation. Science. 272:54–60. - PubMed
-
- Kaech, S.M., E.J. Wherry, and R. Ahmed. 2002. Effector and memory T-cell differentiation: implications for vaccine development. Nat. Rev. Immunol. 2:251–262. - PubMed
-
- Mercado, R., S. Vijh, S.E. Allen, K. Kerksiek, I.M. Pilip, and E.G. Pamer. 2000. Early programming of T cell populations responding to bacterial infection. J. Immunol. 165:6833–6839. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
