

Structures of protein domains that create or recognize histone modifications
- PMID: 15184976
- PMCID: PMC1299057
- DOI: 10.1038/sj.embor.7400146
Structures of protein domains that create or recognize histone modifications
Abstract
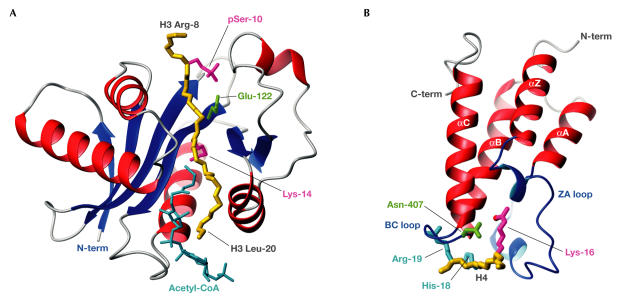
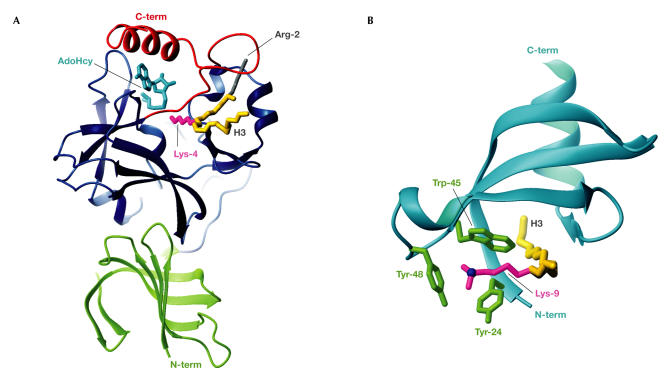
DNA is packed together with histone proteins in cell nuclei to form a compact structure called chromatin. Chromatin represents a scaffold for many genetic events and shows varying degrees of condensation, including a relatively open form (euchromatin) and a highly condensed form (heterochromatin). Enzymes such as histone acetyltransferases (HATs) and methylases covalently label the amino-termini of histones, thereby creating a 'histone code' of modifications that is interpreted by the recruitment of other proteins through recognition domains. Ultimately, this network of interacting proteins is thought to control the degree of chromatin condensation so that DNA is available when it is required for genomic processes. Reviewed here are the structures of HAT and SET domains, which mediate the acetylation and methylation of histones, respectively, and bromodomains and chromodomains, which recognize the modified histones. How these structures have increased our understanding of DNA regulation is also discussed.
Figures


References
-
- Akhtar A, Zink D, Becker PB (2000) Chromodomains are protein–RNA interaction modules. Nature 407: 405–409 - PubMed
-
- Bottomley MJ, Collard MW, Huggenvik JI, Liu Z, Gibson TJ, Sattler M (2001) The SAND domain structure defines a novel DNA-binding fold in transcriptional regulation. Nat Struct Biol 8: 626–633 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
