

Compartmentalization of gene expression during Bacillus subtilis spore formation
- PMID: 15187183
- PMCID: PMC419919
- DOI: 10.1128/MMBR.68.2.234-262.2004
Compartmentalization of gene expression during Bacillus subtilis spore formation
Abstract
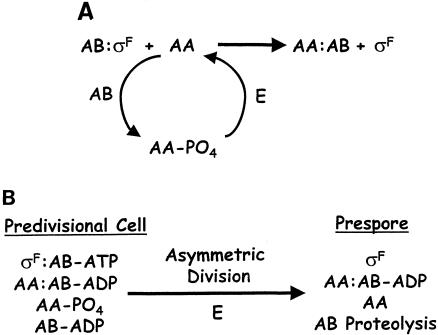
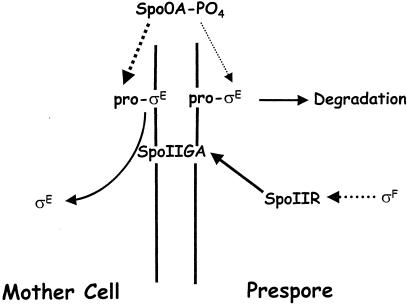
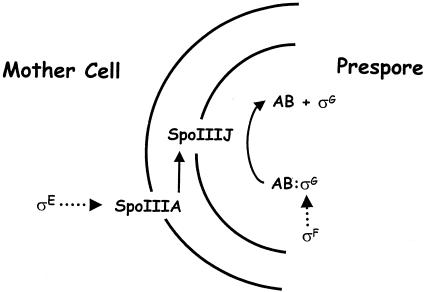
Gene expression in members of the family Bacillaceae becomes compartmentalized after the distinctive, asymmetrically located sporulation division. It involves complete compartmentalization of the activities of sporulation-specific sigma factors, sigma(F) in the prespore and then sigma(E) in the mother cell, and then later, following engulfment, sigma(G) in the prespore and then sigma(K) in the mother cell. The coupling of the activation of sigma(F) to septation and sigma(G) to engulfment is clear; the mechanisms are not. The sigma factors provide the bare framework of compartment-specific gene expression. Within each sigma regulon are several temporal classes of genes, and for key regulators, timing is critical. There are also complex intercompartmental regulatory signals. The determinants for sigma(F) regulation are assembled before septation, but activation follows septation. Reversal of the anti-sigma(F) activity of SpoIIAB is critical. Only the origin-proximal 30% of a chromosome is present in the prespore when first formed; it takes approximately 15 min for the rest to be transferred. This transient genetic asymmetry is important for prespore-specific sigma(F) activation. Activation of sigma(E) requires sigma(F) activity and occurs by cleavage of a prosequence. It must occur rapidly to prevent the formation of a second septum. sigma(G) is formed only in the prespore. SpoIIAB can block sigma(G) activity, but SpoIIAB control does not explain why sigma(G) is activated only after engulfment. There is mother cell-specific excision of an insertion element in sigK and sigma(E)-directed transcription of sigK, which encodes pro-sigma(K). Activation requires removal of the prosequence following a sigma(G)-directed signal from the prespore.
Figures




Similar articles
-
Blocking chromosome translocation during sporulation of Bacillus subtilis can result in prespore-specific activation of sigmaG that is independent of sigmaE and of engulfment.J Bacteriol. 2006 Oct;188(20):7267-73. doi: 10.1128/JB.00744-06. J Bacteriol. 2006. PMID: 17015665 Free PMC article.
-
Control of the expression and compartmentalization of (sigma)G activity during sporulation of Bacillus subtilis by regulators of (sigma)F and (sigma)E.J Bacteriol. 2005 Oct;187(19):6832-40. doi: 10.1128/JB.187.19.6832-6840.2005. J Bacteriol. 2005. PMID: 16166546 Free PMC article.
-
Expression of spoIIIJ in the prespore is sufficient for activation of sigma G and for sporulation in Bacillus subtilis.J Bacteriol. 2003 Jul;185(13):3905-17. doi: 10.1128/JB.185.13.3905-3917.2003. J Bacteriol. 2003. PMID: 12813085 Free PMC article.
-
Control of the cell-specificity of sigma F activity in Bacillus subtilis.Philos Trans R Soc Lond B Biol Sci. 1996 Apr 29;351(1339):537-42. doi: 10.1098/rstb.1996.0052. Philos Trans R Soc Lond B Biol Sci. 1996. PMID: 8735276 Review.
-
Differential gene expression in genetically identical sister cells: the initiation of sporulation in Bacillus subtilis.Mol Microbiol. 2005 May;56(3):578-89. doi: 10.1111/j.1365-2958.2005.04594.x. Mol Microbiol. 2005. PMID: 15819616 Review.
Cited by
-
How Quorum Sensing Connects Sporulation to Necrotrophism in Bacillus thuringiensis.PLoS Pathog. 2016 Aug 2;12(8):e1005779. doi: 10.1371/journal.ppat.1005779. eCollection 2016 Aug. PLoS Pathog. 2016. PMID: 27483473 Free PMC article.
-
Adaptive strategies and pathogenesis of Clostridium difficile from in vivo transcriptomics.Infect Immun. 2013 Oct;81(10):3757-69. doi: 10.1128/IAI.00515-13. Epub 2013 Jul 29. Infect Immun. 2013. PMID: 23897605 Free PMC article.
-
The LysR-type transcriptional regulator YofA controls cell division through the regulation of expression of ftsW in Bacillus subtilis.J Bacteriol. 2007 Aug;189(15):5642-51. doi: 10.1128/JB.00467-07. Epub 2007 May 25. J Bacteriol. 2007. PMID: 17526699 Free PMC article.
-
The Bacillus subtilis DivIVA protein has a sporulation-specific proximity to Spo0J.J Bacteriol. 2006 Aug;188(16):6039-43. doi: 10.1128/JB.01750-05. J Bacteriol. 2006. PMID: 16885474 Free PMC article.
-
Cytological analysis of the mother cell death process during sporulation in Bacillus subtilis.J Bacteriol. 2007 Mar;189(6):2561-5. doi: 10.1128/JB.01738-06. Epub 2007 Jan 5. J Bacteriol. 2007. PMID: 17209033 Free PMC article.
References
-
- Alper, S., L. Duncan, and R. Losick. 1994. An adenosine nucleotide switch controlling the activity of a cell type-specific transcription factor in B. subtilis. Cell 77:195-205. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
