

Functional unit of the RNA polymerase II C-terminal domain lies within heptapeptide pairs
- PMID: 15189994
- PMCID: PMC420137
- DOI: 10.1128/EC.3.3.735-740.2004
Functional unit of the RNA polymerase II C-terminal domain lies within heptapeptide pairs
Abstract
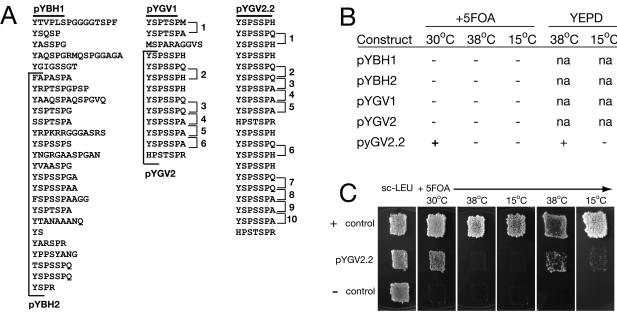
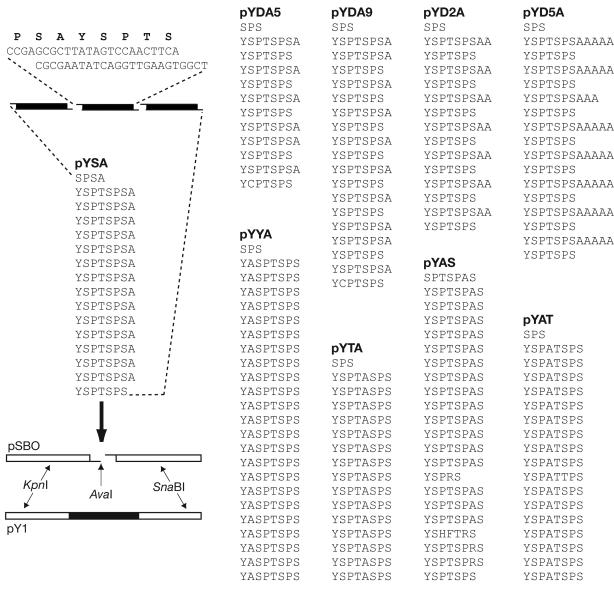
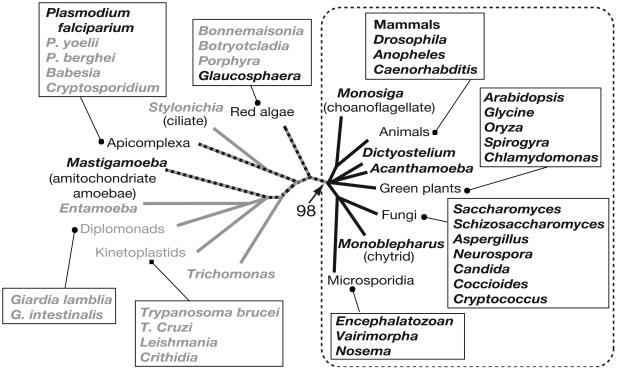
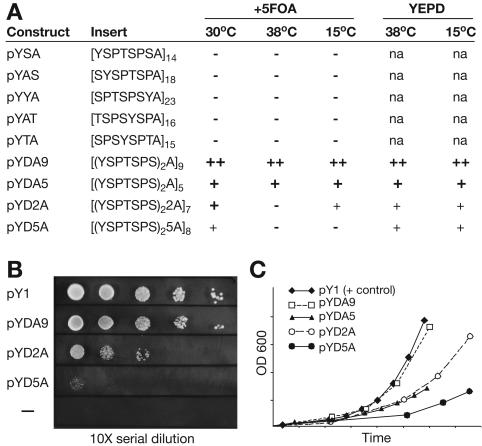
Unlike all other RNA polymerases, the largest subunit (RPB1) of eukaryotic DNA-dependent RNA polymerase II (RNAP II) has a C-terminal domain (CTD) comprising tandemly repeated heptapeptides with the consensus sequence Y-S-P-T-S-P-S. The tandem structure, heptad consensus, and most key functions of the CTD are conserved between yeast and mammals. In fact, all metazoans, fungi, and green plants examined to date, as well as the nearest protistan relatives of these multicellular groups, contain a tandemly repeated CTD. In contrast, the RNAP II largest subunits from many other eukaryotic organisms have a highly degenerate C terminus or show no semblance of the CTD whatsoever. The reasons for intense stabilizing selection on CTD structure in certain eukaryotes, and its apparent absence in others, are unknown. Here we demonstrate, through in vivo genetic complementation, that the essential functional unit of the yeast CTD is contained within pairs of heptapeptides. Insertion of a single alanine residue between diheptads has little phenotypic effect, while increasing the distance between diheptads produces a mostly quantitative effect on yeast cell growth. We further explore structural constraints on the CTD within an evolutionary context and propose selective mechanisms that could maintain a global tandem structure across hundreds of millions of years of eukaryotic evolution.
Copyright 2004 American Society for Microbiology
Figures
References
-
- Allison, L. A., J. K. Wong, V. D. Fitzpatrick, M. Moyle, and C. J. Ingles. 1988. The C-terminal domain of the largest subunit of RNA polymerase II of Saccharomyces cerevisiae, Drosophila melanogaster, and mammals: a conserved structure with an essential function. Mol. Cell. Biol. 8:321-329. - PMC - PubMed
-
- Boeke, J. D., J. Trueheart, G. Natsoulis, and G. R. Fink. 1987. 5-Fluoro-orotic acid as a selective agent in yeast molecular genetics. Methods Enzymol. 154:164-175. - PubMed
-
- Calvo, O., and J. L. Manley. 2003. Strange bedfellows: polyadenlyation factors at the promoter. Genes Dev. 17:1321-1327. - PubMed
-
- Carty, S. M., and A. L. Greenleaf. 2002. Phospho- CTD-association proteins in the nuclear proteome link transcription to DNA/chromatin modification and RNA procession. Mol. Cell. Proteomics 1:598-610. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
