

Activation of alternative NF-kappa B pathway by human herpes virus 8-encoded Fas-associated death domain-like IL-1 beta-converting enzyme inhibitory protein (vFLIP)
- PMID: 15190178
- PMCID: PMC438988
- DOI: 10.1073/pnas.0308016101
Activation of alternative NF-kappa B pathway by human herpes virus 8-encoded Fas-associated death domain-like IL-1 beta-converting enzyme inhibitory protein (vFLIP)
Abstract
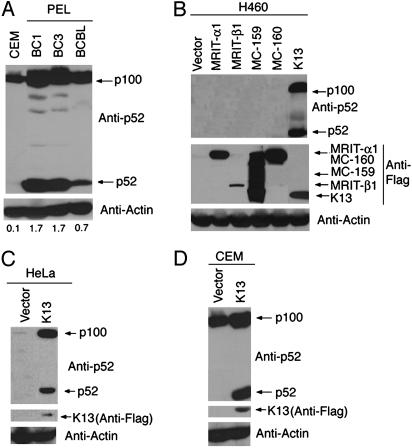
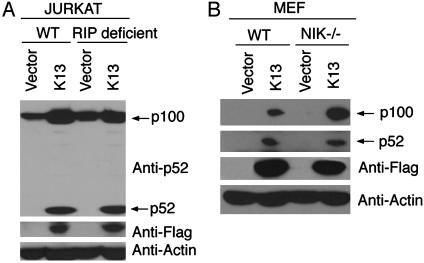
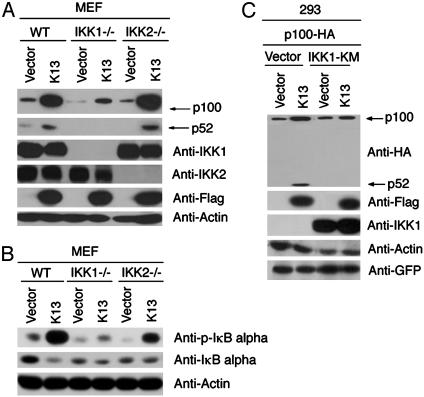
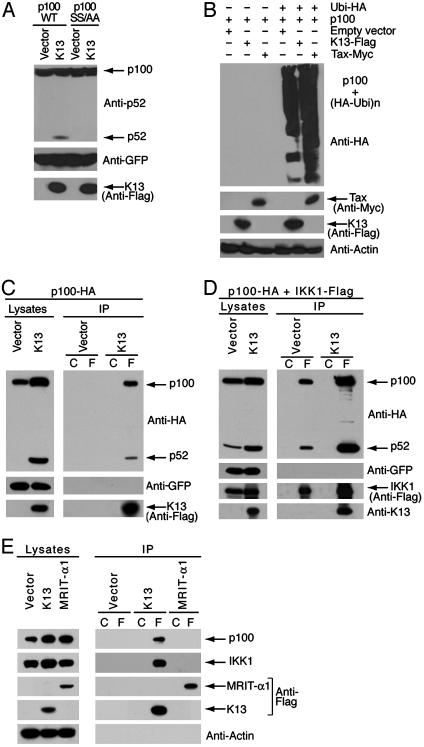
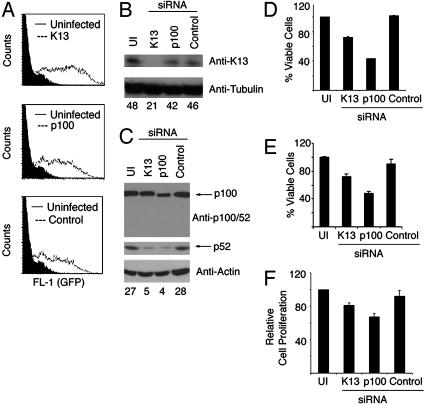
The Kaposi's sarcoma-associated herpesvirus (KSHV, also called human herpesvirus 8) has been linked to KS and primary effusion lymphoma (PEL) in immunocompromised individuals. We report that PEL cell lines have constitutive active alternative NF-kappa B pathway and demonstrate high-level expression of NF-kappa B2/p100 precursor and its processed subunit p52. To elucidate the mechanism of activation of the alternative NF-kappa B pathway in PEL cells, we have investigated the role of KSHV-encoded viral Fas-associated death domain-like IL- beta 1-converting enzyme inhibitory protein (vFLIP) K13. We demonstrate that stable expression of K13, but not other FLIPs, in a variety of cell lines constitutively up-regulates p100/NF-kappa B2 expression and leads to its processing into the p52 subunit. K13-induced up-regulation and processing of p100 critically depends on the I kappa B kinase (IKK)alpha/IKK1 subunit of the IKK complex, whereas IKK beta/IKK2, receptor-interacting protein, and NF-kappa B-inducing kinase are dispensable for this process. Silencing of endogenous K13 expression by siRNA inhibits p100 processing and cellular proliferation. Our results demonstrate for the first time, to our knowledge, that KSHV vFLIP K13 is required for the growth and proliferation of PEL cells and alternative NF-kappa B pathway plays a key role in this process. Therapeutic agents targeting the alternative NF-kappa B pathway may have a role in the treatment of KSHV-associated lymphomas.
Figures
Similar articles
-
Constitutive NF-kappaB activation, normal Fas-induced apoptosis, and increased incidence of lymphoma in human herpes virus 8 K13 transgenic mice.Proc Natl Acad Sci U S A. 2005 Sep 6;102(36):12885-90. doi: 10.1073/pnas.0408577102. Epub 2005 Aug 24. Proc Natl Acad Sci U S A. 2005. PMID: 16120683 Free PMC article.
-
Molecular genetic analysis of human herpes virus 8-encoded viral FLICE inhibitory protein-induced NF-kappaB activation.J Biol Chem. 2003 Dec 26;278(52):52406-11. doi: 10.1074/jbc.M307308200. Epub 2003 Oct 15. J Biol Chem. 2003. PMID: 14561765
-
Nm23-H1 induces apoptosis in primary effusion lymphoma cells via inhibition of NF-κB signaling through interaction with oncogenic latent protein vFLIP K13 of Kaposi's sarcoma-associated herpes virus.Cell Oncol (Dordr). 2022 Oct;45(5):967-989. doi: 10.1007/s13402-022-00701-9. Epub 2022 Aug 14. Cell Oncol (Dordr). 2022. PMID: 35964258
-
Kaposi's sarcoma associated herpesvirus encoded viral FLICE inhibitory protein K13 activates NF-κB pathway independent of TRAF6, TAK1 and LUBAC.PLoS One. 2012;7(5):e36601. doi: 10.1371/journal.pone.0036601. Epub 2012 May 8. PLoS One. 2012. PMID: 22590573 Free PMC article.
-
Regulation of the MIR155 host gene in physiological and pathological processes.Gene. 2013 Dec 10;532(1):1-12. doi: 10.1016/j.gene.2012.12.009. Epub 2012 Dec 14. Gene. 2013. PMID: 23246696 Review.
Cited by
-
Constitutive NF-kappaB activation, normal Fas-induced apoptosis, and increased incidence of lymphoma in human herpes virus 8 K13 transgenic mice.Proc Natl Acad Sci U S A. 2005 Sep 6;102(36):12885-90. doi: 10.1073/pnas.0408577102. Epub 2005 Aug 24. Proc Natl Acad Sci U S A. 2005. PMID: 16120683 Free PMC article.
-
A20 is induced by Kaposi sarcoma-associated herpesvirus-encoded viral FLICE inhibitory protein (vFLIP) K13 and blocks K13-induced nuclear factor-kappaB in a negative feedback manner.J Biol Chem. 2011 Jun 17;286(24):21555-64. doi: 10.1074/jbc.M111.224048. Epub 2011 Apr 29. J Biol Chem. 2011. PMID: 21531730 Free PMC article.
-
Emmprin and KSHV: new partners in viral cancer pathogenesis.Cancer Lett. 2013 Sep 1;337(2):161-6. doi: 10.1016/j.canlet.2013.05.037. Epub 2013 Jun 3. Cancer Lett. 2013. PMID: 23743354 Free PMC article. Review.
-
Kaposi's Sarcoma-Associated Herpesvirus Genome Replication, Partitioning, and Maintenance in Latency.Front Microbiol. 2012 Jan 24;3:7. doi: 10.3389/fmicb.2012.00007. eCollection 2012. Front Microbiol. 2012. PMID: 22291692 Free PMC article.
-
NEMO is essential for Kaposi's sarcoma-associated herpesvirus-encoded vFLIP K13-induced gene expression and protection against death receptor-induced cell death, and its N-terminal 251 residues are sufficient for this process.J Virol. 2014 Jun;88(11):6345-54. doi: 10.1128/JVI.00028-14. Epub 2014 Mar 26. J Virol. 2014. PMID: 24672029 Free PMC article.
References
-
- Thome, M., Schneider, P., Hofmann, K., Fickenscher, H., Meinl, E., Neipel, F., Mattmann, C., Burns, K., Bodmer, J. L., Schroter, M., et al. (1997) Nature 386, 517-521. - PubMed
-
- Chaudhary, P. M., Jasmin, A., Eby, M. T. & Hood, L. (1999) Oncogene 18, 5738-5746. - PubMed
-
- Liu, L., Eby, M. T., Rathore, N., Sinha, S. K., Kumar, A. & Chaudhary, P. M. (2002) J. Biol. Chem. 277, 13745-13751. - PubMed
-
- Field, N., Low, W., Daniels, M., Howell, S., Daviet, L., Boshoff, C. & Collins, M. (2003) J. Cell Sci. 116, 3721-3728. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
