

Differential input by Ste5 scaffold and Msg5 phosphatase route a MAPK cascade to multiple outcomes
- PMID: 15192700
- PMCID: PMC449765
- DOI: 10.1038/sj.emboj.7600250
Differential input by Ste5 scaffold and Msg5 phosphatase route a MAPK cascade to multiple outcomes
Abstract
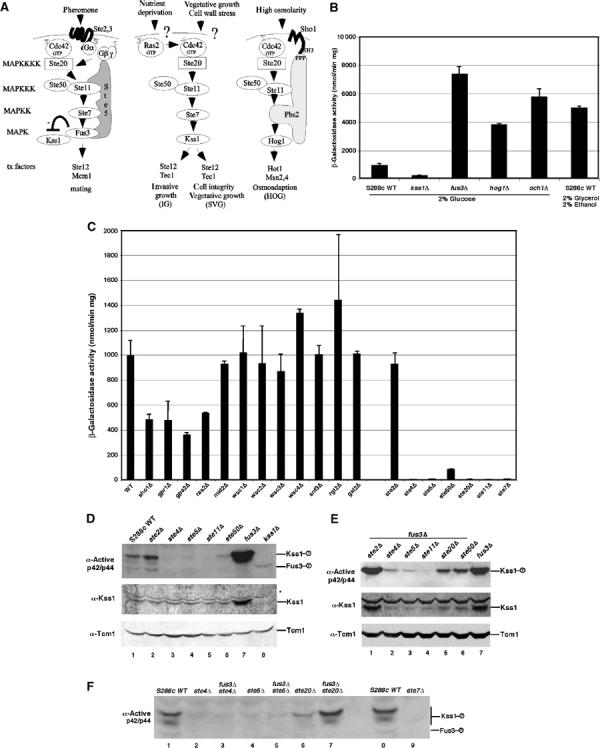
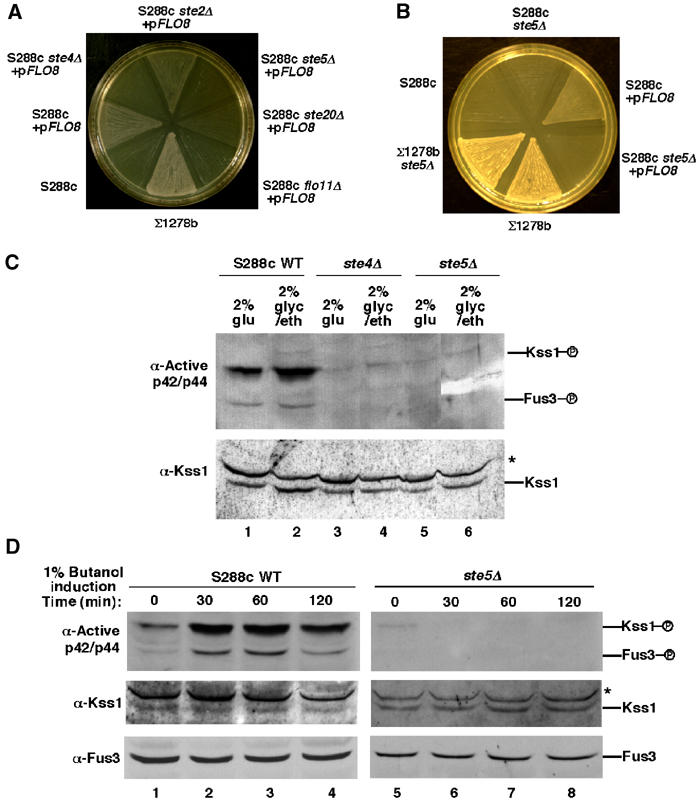
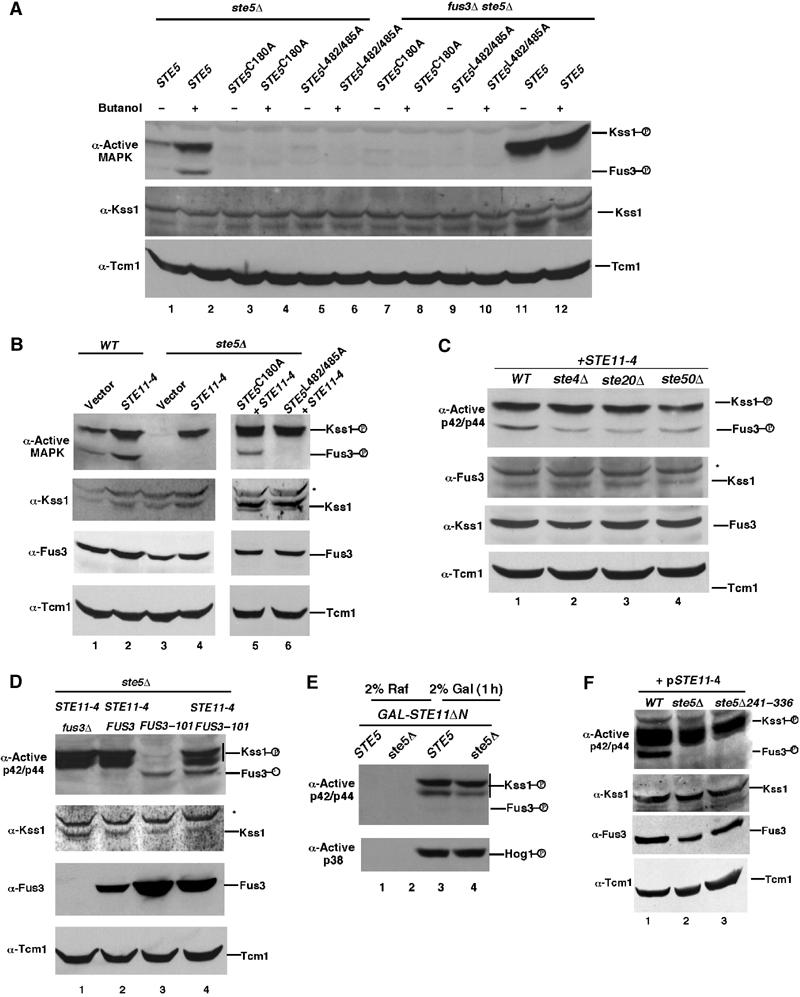
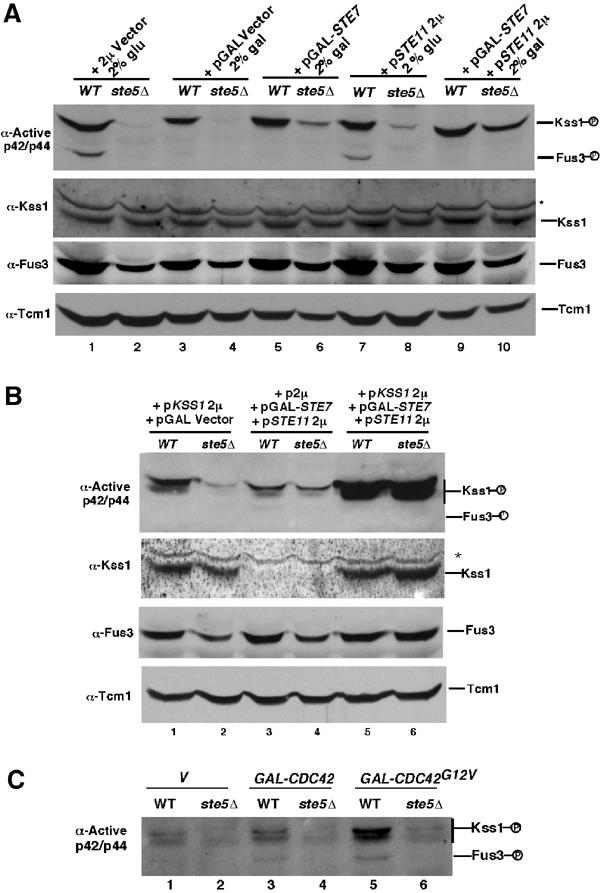
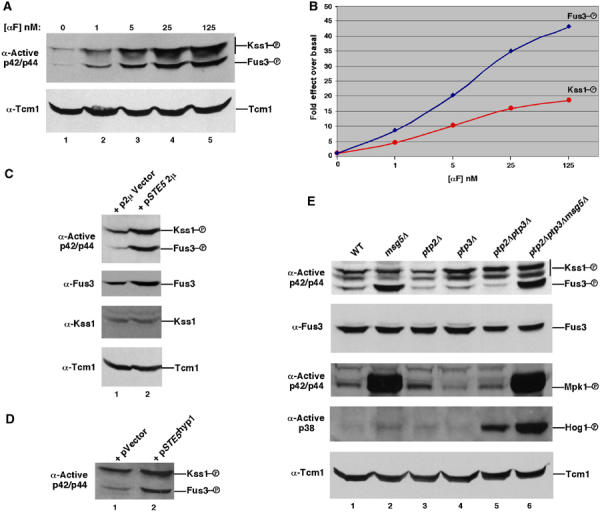
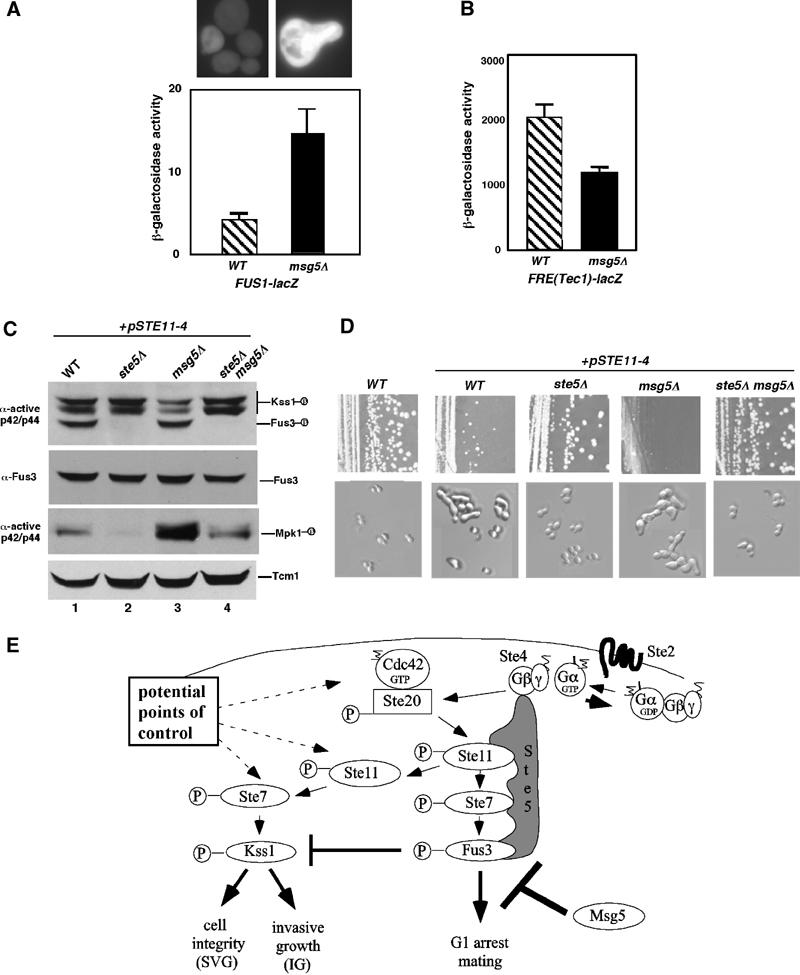
Pathway specificity is poorly understood for mitogen-activated protein kinase (MAPK) cascades that control different outputs in response to different stimuli. In yeast, it is not known how the same MAPK cascade activates Kss1 MAPK to promote invasive growth (IG) and proliferation, and both Fus3 and Kss1 MAPKs to promote mating. Previous work has suggested that the Kss1 MAPK cascade is activated independently of the mating G protein (Ste4)-scaffold (Ste5) system during IG. Here we demonstrate that Ste4 and Ste5 activate Kss1 during IG and in response to multiple stimuli including butanol. Ste5 activates Kss1 by generating a pool of active MAPKKK (Ste11), whereas additional scaffolding is needed to activate Fus3. Scaffold-independent activation of Kss1 can occur at multiple steps in the pathway, whereas Fus3 is strictly dependent on the scaffold. Pathway specificity is linked to Kss1 immunity to a MAPK phosphatase that constitutively inhibits basal activation of Fus3 and blocks activation of the mating pathway. These findings reveal the versatility of scaffolds and how a single MAPK cascade mediates different outputs.
Figures
Similar articles
-
Persistent activation by constitutive Ste7 promotes Kss1-mediated invasive growth but fails to support Fus3-dependent mating in yeast.Mol Cell Biol. 2004 Oct;24(20):9221-38. doi: 10.1128/MCB.24.20.9221-9238.2004. Mol Cell Biol. 2004. PMID: 15456892 Free PMC article.
-
Effects of HSP70 chaperones Ssa1 and Ssa2 on Ste5 scaffold and the mating mitogen-activated protein kinase (MAPK) pathway in Saccharomyces cerevisiae.PLoS One. 2023 Oct 18;18(10):e0289339. doi: 10.1371/journal.pone.0289339. eCollection 2023. PLoS One. 2023. PMID: 37851593 Free PMC article.
-
MAPK specificity in the yeast pheromone response independent of transcriptional activation.Curr Biol. 2001 Aug 21;11(16):1266-71. doi: 10.1016/s0960-9822(01)00370-0. Curr Biol. 2001. PMID: 11525741
-
Mitogen-activated protein kinases with distinct requirements for Ste5 scaffolding influence signaling specificity in Saccharomyces cerevisiae.Mol Cell Biol. 2005 Mar;25(5):1793-803. doi: 10.1128/MCB.25.5.1793-1803.2005. Mol Cell Biol. 2005. PMID: 15713635 Free PMC article.
-
Pheromone response, mating and cell biology.Curr Opin Microbiol. 2000 Dec;3(6):573-81. doi: 10.1016/s1369-5274(00)00143-0. Curr Opin Microbiol. 2000. PMID: 11121776 Review.
Cited by
-
Persistent activation by constitutive Ste7 promotes Kss1-mediated invasive growth but fails to support Fus3-dependent mating in yeast.Mol Cell Biol. 2004 Oct;24(20):9221-38. doi: 10.1128/MCB.24.20.9221-9238.2004. Mol Cell Biol. 2004. PMID: 15456892 Free PMC article.
-
Control of MAPK signaling specificity by a conserved residue in the MEK-binding domain of the yeast scaffold protein Ste5.Curr Genet. 2006 Jun;49(6):351-63. doi: 10.1007/s00294-006-0061-6. Epub 2006 Feb 4. Curr Genet. 2006. PMID: 16463042
-
Mechanisms of MAPK signalling specificity.Biochem Soc Trans. 2006 Nov;34(Pt 5):837-41. doi: 10.1042/BST0340837. Biochem Soc Trans. 2006. PMID: 17052210 Free PMC article.
-
Glycosylation defects activate filamentous growth Kss1 MAPK and inhibit osmoregulatory Hog1 MAPK.EMBO J. 2009 May 20;28(10):1380-91. doi: 10.1038/emboj.2009.104. Epub 2009 Apr 16. EMBO J. 2009. PMID: 19369942 Free PMC article.
-
Regulation of cell signaling dynamics by the protein kinase-scaffold Ste5.Mol Cell. 2008 Jun 6;30(5):649-56. doi: 10.1016/j.molcel.2008.04.016. Mol Cell. 2008. PMID: 18538663 Free PMC article.
References
-
- Bhalla US, Ram PT, Iyengar R (2002) MAP kinase phosphatase as a locus of flexibility in a mitogen-activated protein kinase signaling network. Science 297: 1018–1023 - PubMed
-
- Breitkreutz A, Tyers M (2002) MAPK signaling specificity: it takes two to tango. Trends Cell Biol 12: 254–257 - PubMed
-
- Breitkreutz A, Boucher L, tyers M. (2001) MAPK specificity in the yeast pheromone response independent of transcriptional activation. Curr Biol 11: 1266–1271 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
