

PS1 activates PI3K thus inhibiting GSK-3 activity and tau overphosphorylation: effects of FAD mutations
- PMID: 15192701
- PMCID: PMC449766
- DOI: 10.1038/sj.emboj.7600251
PS1 activates PI3K thus inhibiting GSK-3 activity and tau overphosphorylation: effects of FAD mutations
Abstract
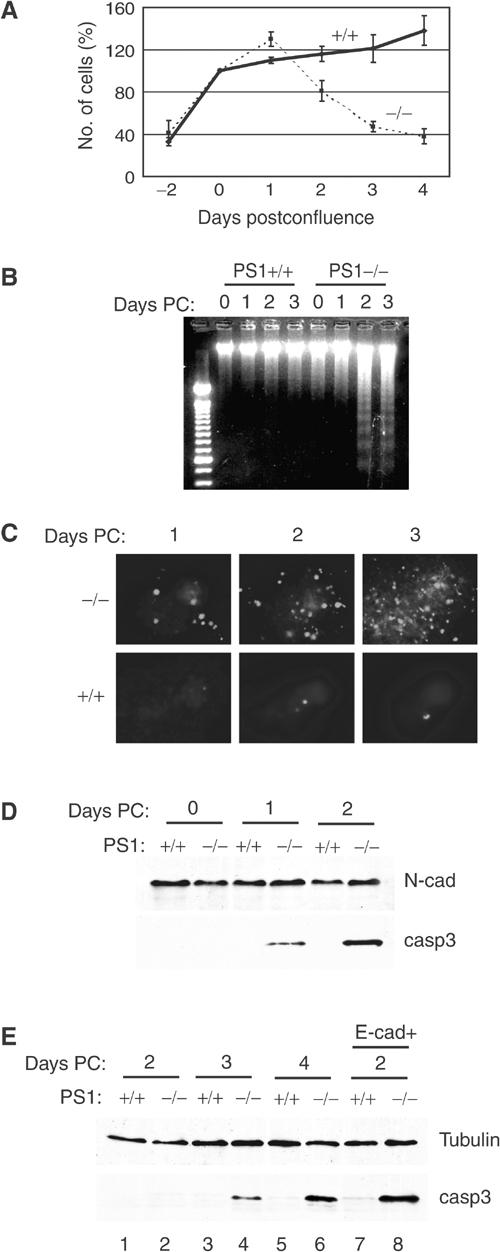
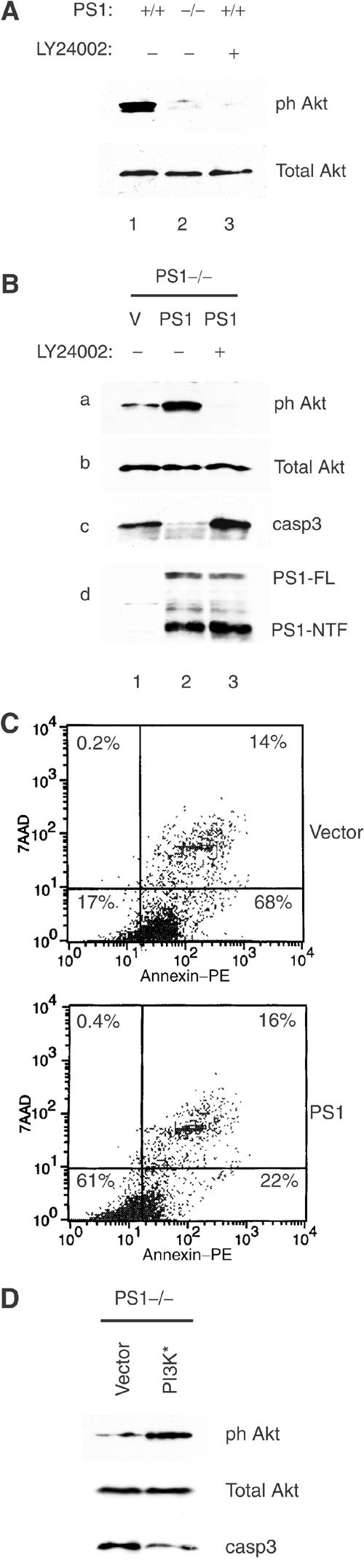
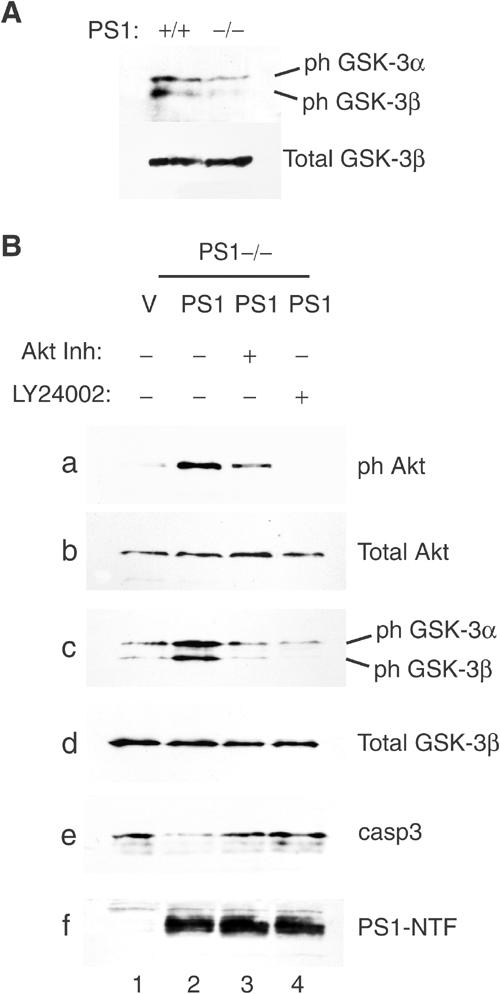
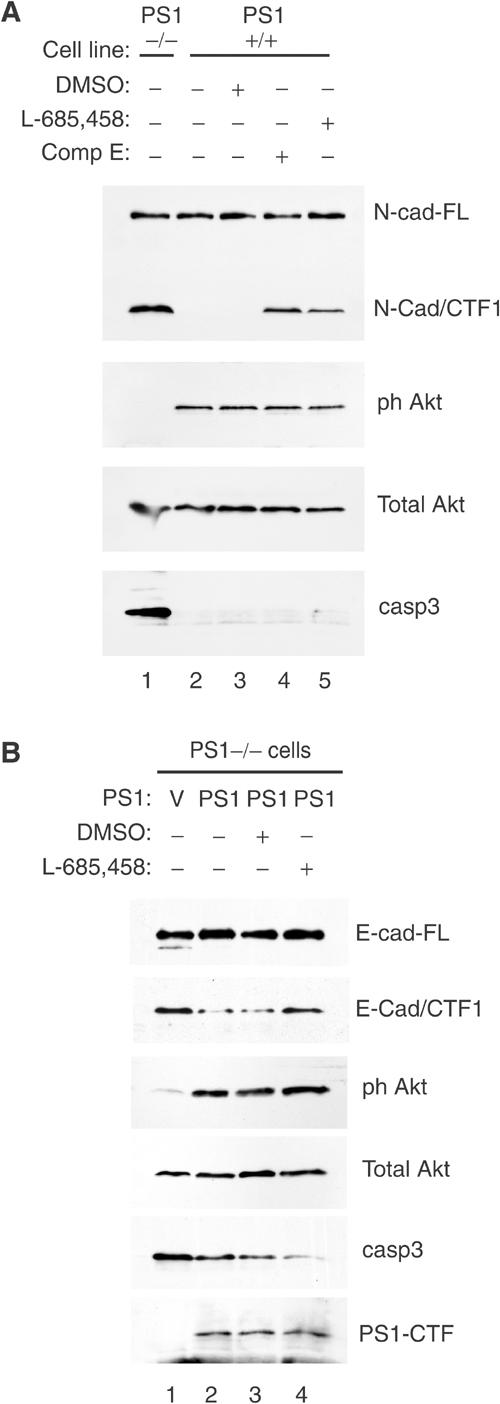
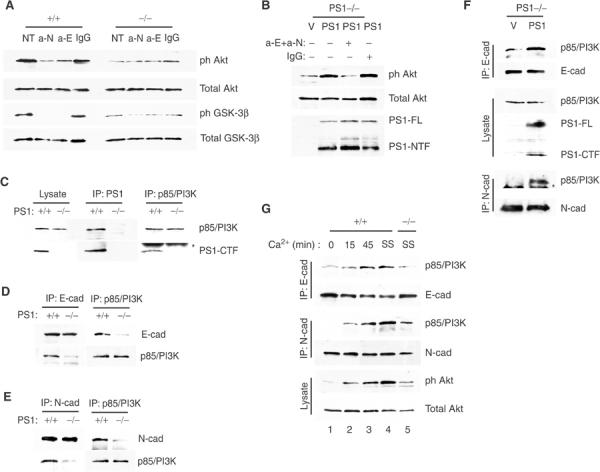
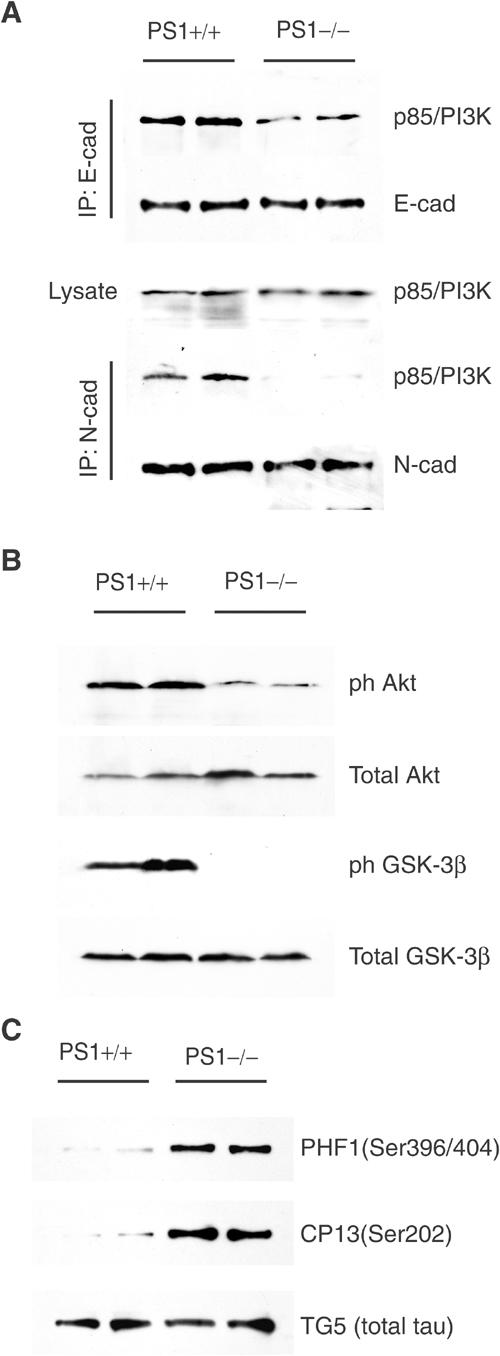
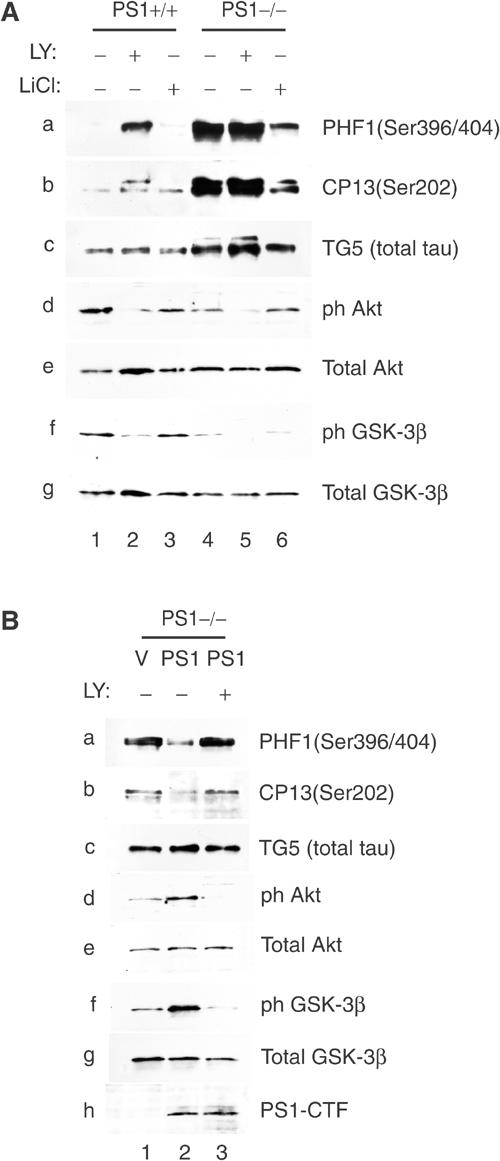
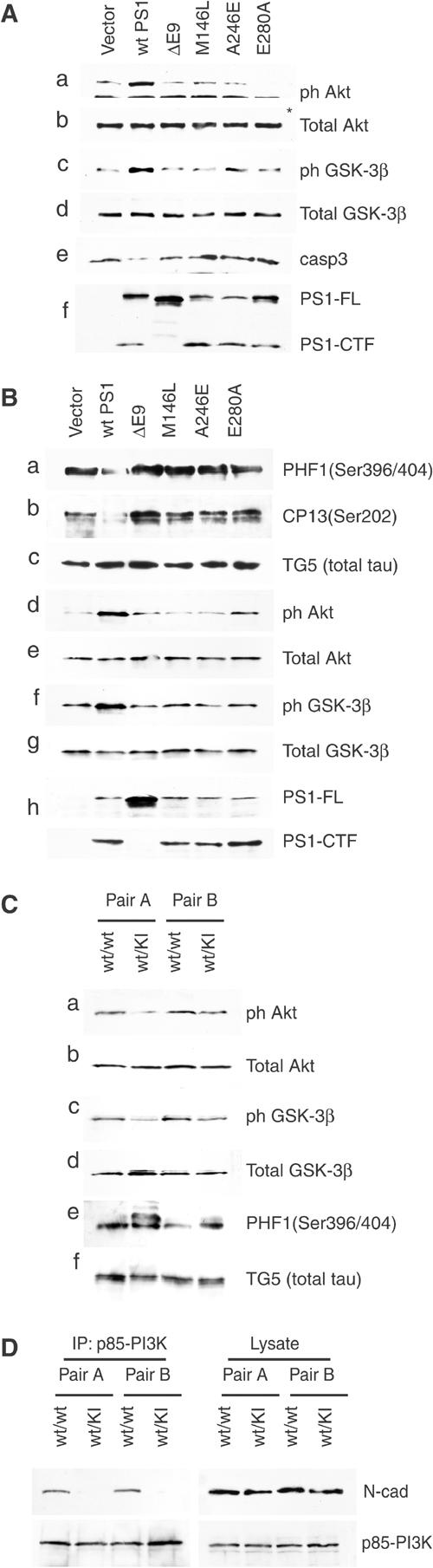
Phosphatidylinositol 3-kinase (PI3K) promotes cell survival and communication by activating its downstream effector Akt kinase. Here we show that PS1, a protein involved in familial Alzheimer's disease (FAD), promotes cell survival by activating the PI3K/Akt cell survival signaling. This function of PS1 is unaffected by gamma-secretase inhibitors. Pharmacological and genetic evidence indicates that PS1 acts upstream of Akt, at or before PI3K kinase. PS1 forms complexes with the p85 subunit of PI3K and promotes cadherin/PI3K association. Furthermore, conditions that inhibit this association prevent the PS1-induced PI3K/Akt activation, indicating that PS1 stimulates PI3K/Akt signaling by promoting cadherin/PI3K association. By activating PI3K/Akt signaling, PS1 promotes phosphorylation/inactivation of glycogen synthase kinase-3 (GSK-3), suppresses GSK-3-dependent phosphorylation of tau at residues overphosphorylated in AD and prevents apoptosis of confluent cells. PS1 FAD mutations inhibit the PS1-dependent PI3K/Akt activation, thus promoting GSK-3 activity and tau overphosphorylation at AD-related residues. Our data raise the possibility that PS1 may prevent development of AD pathology by activating the PI3K/Akt signaling pathway. In contrast, FAD mutations may promote AD pathology by inhibiting this pathway.
Figures
References
-
- Baki L, Marambaud P, Efthimiopoulos S, Georgakopoulos A, Wen P, Cui W, Shioi J, Koo E, Ozawa M, Friedrich VL Jr, Robakis NK (2001) Presenilin-1 binds cytoplasmic epithelial cadherin, inhibits cadherin/p120 association, and regulates stability and function of the cadherin/catenin adhesion complex. Proc Natl Acad Sci USA 98: 2381–2386 - PMC - PubMed
-
- Brunet A, Datta SR, Greenberg ME (2001) Transcription-dependent and -independent control of neuronal survival by the PI3K–Akt signaling pathway. Curr Opin Neurobiol 11: 297–305 - PubMed
-
- Chan TO, Rittenhouse SE, Tsichlis PN (1999) AKT/PKB and other D3 phosphoinositide-regulated kinases: kinase activation by phosphoinositide-dependent phosphorylation. Annu Rev Biochem 68: 965–1014 - PubMed
-
- Chan TO, Tsichlis PN (2001) PDK2: a complex tail in one Akt. Sci STKE 2001: PE1. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
