

Modulation of the polyamine biosynthetic pathway in transgenic rice confers tolerance to drought stress
- PMID: 15197268
- PMCID: PMC470772
- DOI: 10.1073/pnas.0306974101
Modulation of the polyamine biosynthetic pathway in transgenic rice confers tolerance to drought stress
Abstract
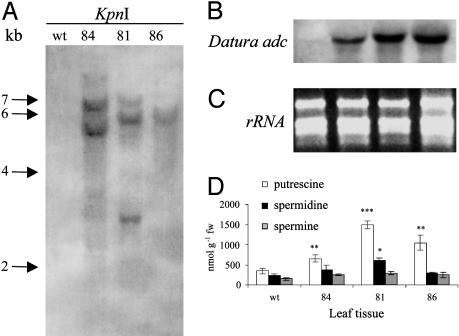
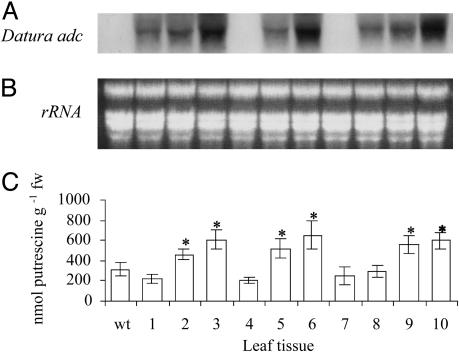
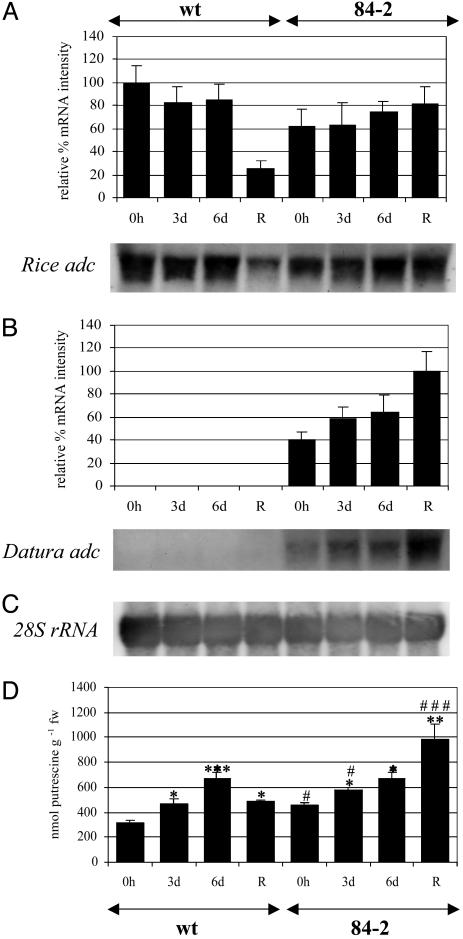
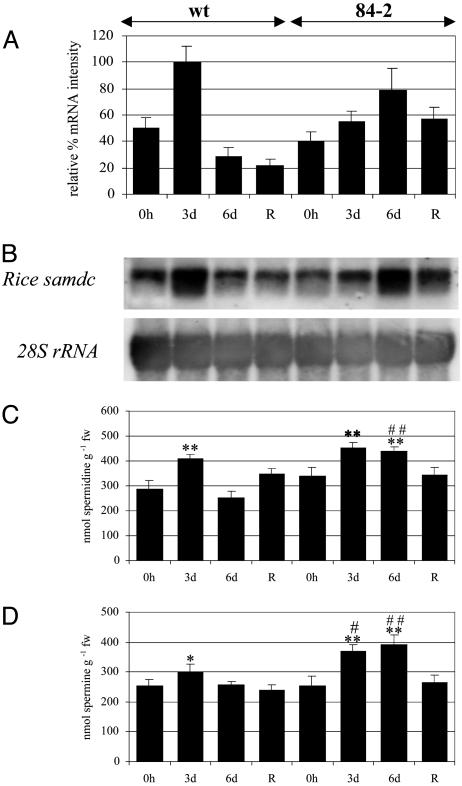
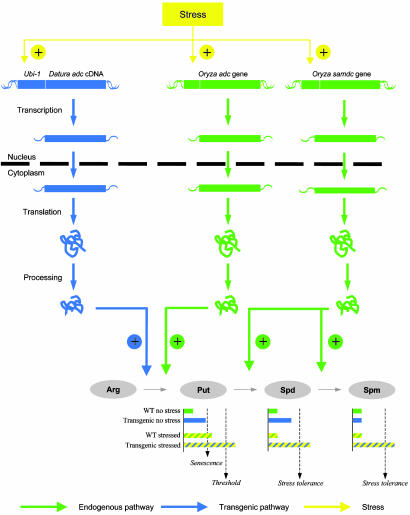
We have generated transgenic rice plants expressing the Datura stramonium adc gene and investigated their response to drought stress. We monitored the steady-state mRNA levels of genes involved in polyamine biosynthesis (Datura adc, rice adc, and rice samdc) and polyamine levels. Wild-type plants responded to the onset of drought stress by increasing endogenous putrescine levels, but this was insufficient to trigger the conversion of putrescine into spermidine and spermine (the agents that are believed to protect plants under stress). In contrast, transgenic plants expressing Datura adc produced much higher levels of putrescine under stress, promoting spermidine and spermine synthesis and ultimately protecting the plants from drought. We demonstrate clearly that the manipulation of polyamine biosynthesis in plants can produce drought-tolerant germplasm, and we propose a model consistent with the role of polyamines in the protection of plants against abiotic stress.
Figures

References
-
- Bajaj, S., Targolli, J., Liu, L. F., Ho, T. H. D. & Wu, R. (1999) Mol. Breeding 5, 493-503.
-
- Richards, F. J. & Coleman, R. G. (1952) Nature 170, 460. - PubMed
-
- Bouchereau, A., Aziz, A., Larher, F. & Martin-Tanguy, J. (1999) Plant Sci. 140, 103-125.
-
- Koening, H., Goldstone, A. D., Lu, C. Y. & Trout, J. J. (1990) Stroke 21, Suppl. III, 98-103. - PubMed
-
- Reggiani, R., Aurisano, N., Mattana, M. & Bertani, A. (1993) J. Plant Physiol. 142, 94-98.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
