

Fossils, feet and the evolution of human bipedal locomotion
- PMID: 15198703
- PMCID: PMC1571304
- DOI: 10.1111/j.0021-8782.2004.00296.x
Fossils, feet and the evolution of human bipedal locomotion
Abstract
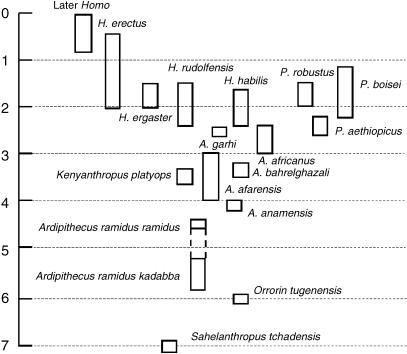
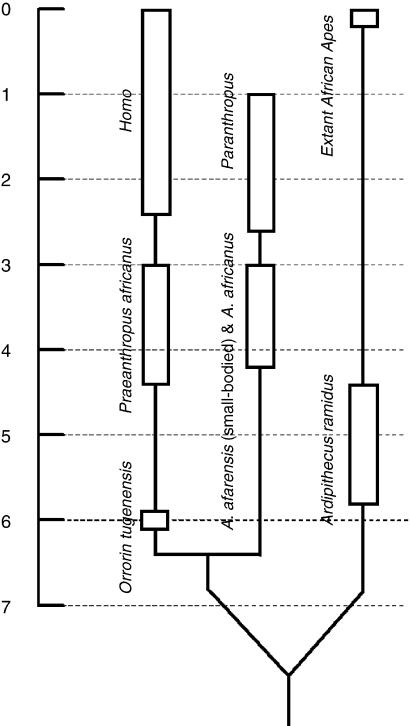
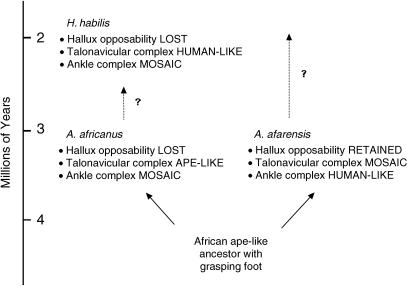
We review the evolution of human bipedal locomotion with a particular emphasis on the evolution of the foot. We begin in the early twentieth century and focus particularly on hypotheses of an ape-like ancestor for humans and human bipedal locomotion put forward by a succession of Gregory, Keith, Morton and Schultz. We give consideration to Morton's (1935) synthesis of foot evolution, in which he argues that the foot of the common ancestor of modern humans and the African apes would be intermediate between the foot of Pan and Hylobates whereas the foot of a hypothetical early hominin would be intermediate between that of a gorilla and a modern human. From this base rooted in comparative anatomy of living primates we trace changing ideas about the evolution of human bipedalism as increasing amounts of postcranial fossil material were discovered. Attention is given to the work of John Napier and John Robinson who were pioneers in the interpretation of Plio-Pleistocene hominin skeletons in the 1960s. This is the period when the wealth of evidence from the southern African australopithecine sites was beginning to be appreciated and Olduvai Gorge was revealing its first evidence for Homo habilis. In more recent years, the discovery of the Laetoli footprint trail, the AL 288-1 (A. afarensis) skeleton, the wealth of postcranial material from Koobi Fora, the Nariokotome Homo ergaster skeleton, Little Foot (Stw 573) from Sterkfontein in South Africa, and more recently tantalizing material assigned to the new and very early taxa Orrorin tugenensis, Ardipithecus ramidus and Sahelanthropus tchadensis has fuelled debate and speculation. The varying interpretations based on this material, together with changing theoretical insights and analytical approaches, is discussed and assessed in the context of new three-dimensional morphometric analyses of australopithecine and Homo foot bones, suggesting that there may have been greater diversity in human bipedalism in the earlier phases of our evolutionary history than previously suspected.
Figures


References
-
- Aiello LC, Dean MC. An Introduction to Human Evolutionary Anatomy. London: Academic Press; 1990.
-
- Aiello LC, Andrews P. The Australopithecines in review. Human Evol. 2000;15:17–38.
-
- Aiello LC, Collard M. Palaeoanthropology: our newest oldest ancestor? Nature. 2001;410:526–527. - PubMed
-
- Asfaw B. Proximal femur articulation in Pliocene hominids. Am. J. Phys Anthropol. 1985;68:535–538. - PubMed
-
- Asfaw B, White T, Lovejoy O, Latimer B, Simpson S, Suwa G. Australopithecus garhi: a new species of early hominid from Ethiopia. Science. 1999;284:629–635. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
