

doi: 10.1101/gad.1199404.
The generation of proper constitutive G-tails on yeast telomeres is dependent on the MRX complex
Affiliations
- PMID: 15198981
- PMCID: PMC423190
- DOI: 10.1101/gad.1199404
Item in Clipboard
The generation of proper constitutive G-tails on yeast telomeres is dependent on the MRX complex
Genes Dev.
.
Abstract
The precise DNA arrangement at chromosomal ends and the proteins involved in its maintenance are of crucial importance for genome stability. For the yeast Saccharomyces cerevisiae, this constitutive DNA configuration has remained unknown. We demonstrate here that G-tails of 12-14 bases are present outside of S phase on normal yeast telomeres. Furthermore, the Mre11p protein is essential for the proper establishment of this constitutive end-structure. However, the timing of extended G-tails occurring during S phase is not affected in strains lacking Mre11p. Thus, G-tails are present on yeast chromosomes throughout the cell cycle and the MRX complex is required for their normal establishment.
Figures
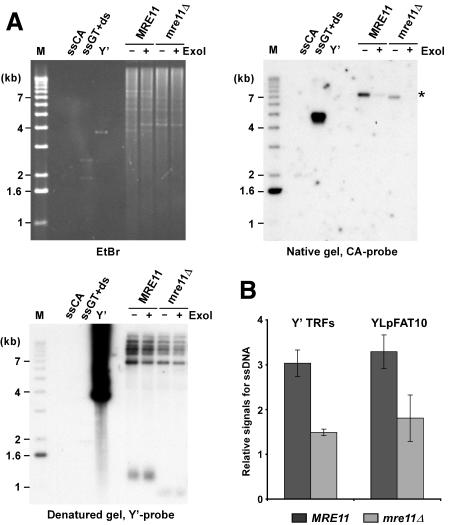
G-tail signals are reduced in mre11Δ strains. (A) Genomic DNA derived from α-factor-arrested cells was digested with XhoI. G-tails on YLpFAT10 (indicated by asterisk) were analyzed by native in-gel hybridization for wild-type and mre11Δ strains (MLY50 and MLY51). DNAs were either mock treated (labeled ExoI-), or treated with Escherichia coli Exonuclease I (labeled ExoI+), before in-gel analysis. (Top, left) Ethidium-bromide-stained gel. (Top, right) The gel after nondenaturing hybridization to the CA probe. (Bottom, left) The DNA was denatured in the gel, transferred to a nylon membrane, and the membrane hybridized to a Y′ probe. Single-stranded phagemid DNA containing yeast telomeric repeats of the G-rich strand (ssGT) and of the C-rich strand (ssCA) served as positive and negative controls, respectively. The ssGT control was mixed with PvuI-digested pMW55, the latter serving as double-stranded control (ds, a fragment of 1.9 kb). Linearized pVZY′K plasmid (3.9 kb) served as a positive control for the hybridization with the Y′ probe (labeled Y′). (M) End-labeled 1-kb ladder DNA serving as a size standard. (B) Relative amounts for G-tails (relative signals for ssDNA, in arbitrary units) were obtained by calculating the ratio of the respective signals on the native gels over the total amount of Y′ TRF signals or YLpFAT10 signals. Three to six individual nonsynchronous cultures were tested for each strain, and the standard deviation is indicated.
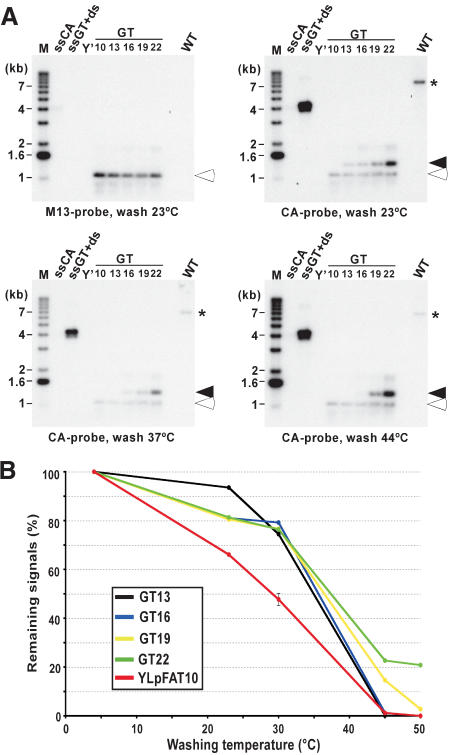
Most G-tails on telomeres in G1-arrested cells are shorter than 16 bases. (A) The individual pGTxx plasmids, each containing telomeric repeat tracts ranging from 10 to 22 bp, were double digested and heat denatured prior to loading on an agarose gel. A common DNA fragment serves as a loading control after hybridization to an M13 probe (open arrowheads), whereas a fragment migrating at ∼1.3 kb of each plasmid contains the TG1–3 repeats of varying lengths serving to compare the hybridization of the CA probe with different telomeric repeat lengths (closed arrowheads). Wild-type cells (MLY30 strain) were arrested in G1 with α-factor, and XhoI-digested DNAs derived from those cells were loaded on the gel (lane marked WT). After hybridization to the M13 probe (top, left), the same gel was then hybridized at 4°C to the CA probe and washed at different temperatures as indicated at the bottom of gels shown. The signals obtained for G-tails on YLpFAT10 (indicated by asterisk) were compared with those detected on the 1.3-kb DNA fragments derived from the pGTxx plasmids after different washing temperatures. Controls and molecular-size standards are as described in Figure 1. (B) Remaining signals on pGTxx fragments, as well as signals for G-tails on YLpFAT10, were determined by using native gels washed at different temperatures. Signals are plotted as percent (%) with respect to the same gel washed at 4°C (designated at 100%).
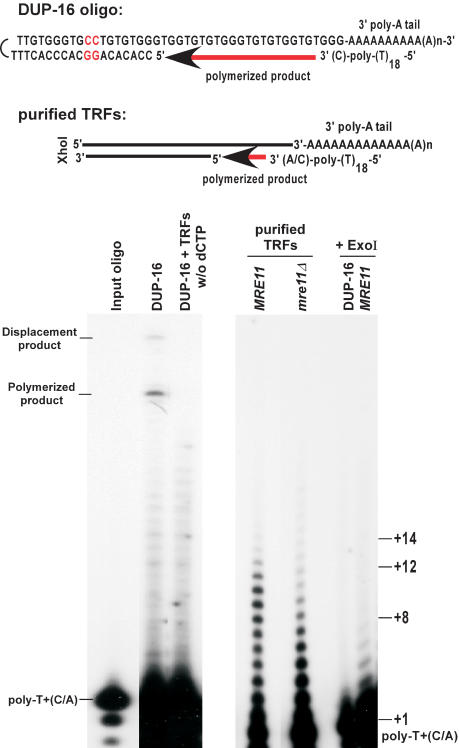
Constitutive G-tails are shorter in mre11Δ cells. (Top) DNA structure of the annealed model substrate DUP-16 and expected structure for TRFs. In DUP-16, the two nucleotides not corresponding to telomeric sequence, in which strand displacement synthesis stops, are indicated in red. (Bottom) Analysis of extension products by PAGE. DUP-16 and/or TRFs were poly(A) tailed, annealed to end-labeled poly-T+(C/A) oligo, and subjected to an extension reaction using T4 DNA polymerase. (Left gel) Control reactions using DUP-16 alone; w/o dCTP; dCTP was omitted from the extension reaction. Poly-T+(C/A) indicates the position of the end-labeled oligo alone, and corresponds to position +1 in terms of overhang size. (Polymerized product) Product of the extension reaction on DUP-16 covering the overhang only (+24); (displacement product) product of the extension reaction reaching the two inversed G:C base pairs (+31). (Right gel) Purified TRFs from the indicated strains were subjected to the procedure. In the last two lanes, either DUP-16 or purified TRFs from a wild-type strain were treated with ExoI before the tailing reaction, and then processed as the other samples. Size indications on right denote overhang size.
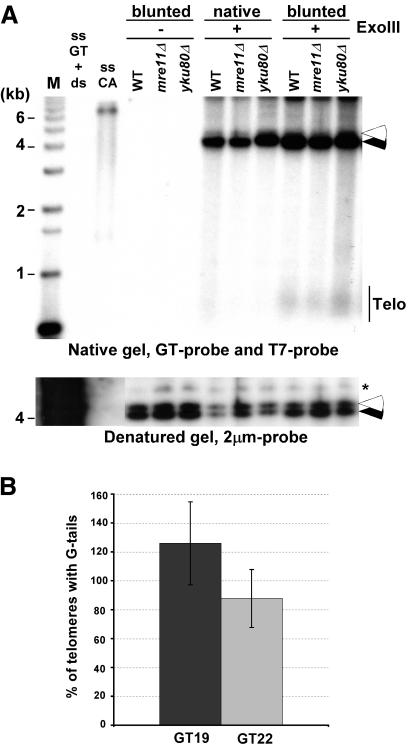
Most yeast telomeres end with a G-tail. (A, top) Genomic DNAs isolated from the indicated strains were first treated with Exonuclease I (labeled blunted) or mock treated (labeled native). DNAs were then mock treated (labeled ExoIII-), or treated with Exonuclease III (labeled ExoIII+) before native gel analysis. (Closed arrowhead) Internal positive control for the ExoIII treatment (Acc65I-digested pRS304); (open arrowhead) internal negative control (KpnI-digested pRS314), revealed by the T7 probe. KpnI-digested TRFs are seen with the GT probe only in lanes in which genomic DNA was blunted with ExoI prior to ExoIII reaction. (Bottom) Same gel as above after denaturation and rehybridization to a 2-μm probe to show equal loading of DNA (indicated by asterisk). Controls and molecular size standards are as in Figure 1. (B) Percentage of telomeres ending with a G-tail was estimated by quantifying the amount of G-tail signals found on YLpFAT10 on native gels washed at 4°C (see Fig. 2A). The values obtained were compared with signals detected for pGT19 and pGT22 fragments and were adjusted for the molecular ratio of hybridization targets on pGT19/pGT22 vs. the YLpFAT10 plasmid (see Supplemental Material). DNAs derived from three independent cultures of wild-type cells were used for quantification, and the standard deviation is indicated.
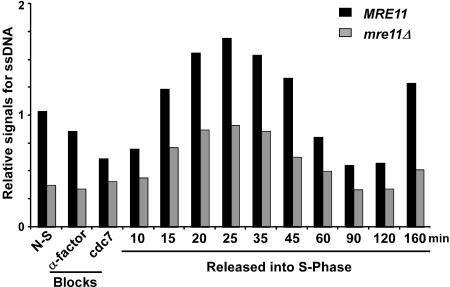
S-phase-dependent G-tail formation in mre11Δ cells. A cell synchronization experiment using MRE11 and mre11Δ cells (AR120 and MLY40 strains) was performed as described in Supplementary Figure 1. DNA isolated from each cell aliquot was analyzed for telomeric DNA end-structures by nondenaturing in-gel hybridization (Supplementary Fig. 2; data not shown). The relative amounts for telomeric ssDNA on YLpFAT10 (in arbitrary units) were obtained by calculating the ratio of the G-tail signals retrieved on the native gel over the total amount of YLpFAT10 detected.
References
-
- Blackburn E.H. 2001. Switching and signaling at the telomere. Cell 106: 661-673. - PubMed
-
- Chakhparonian M. and Wellinger, R.J. 2003. Telomere maintenance and DNA replication: How closely are these two connected? Trends Genet. 19: 439-446. - PubMed
-
- ____. 2002. The Mre11 complex: At the crossroads of dna repair and checkpoint signalling. Nat. Rev. Mol. Cell. Biol. 3: 317-327. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases