

Leukemia proto-oncoprotein MLL forms a SET1-like histone methyltransferase complex with menin to regulate Hox gene expression
- PMID: 15199122
- PMCID: PMC480881
- DOI: 10.1128/MCB.24.13.5639-5649.2004
Leukemia proto-oncoprotein MLL forms a SET1-like histone methyltransferase complex with menin to regulate Hox gene expression
Abstract
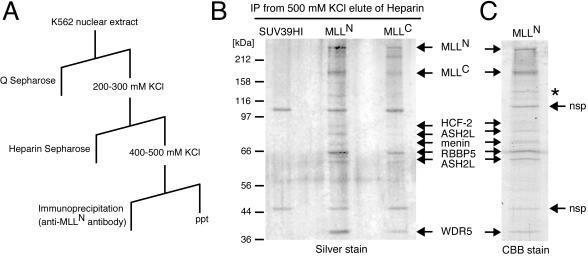
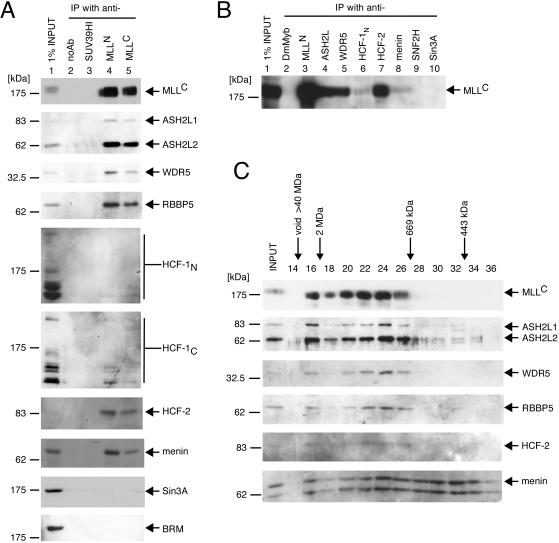
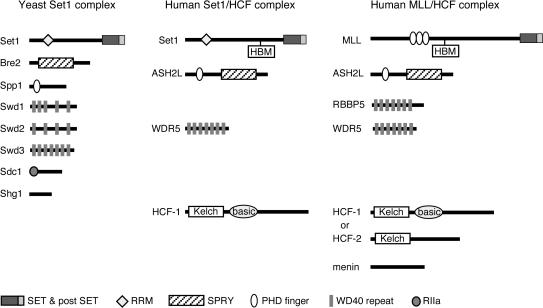
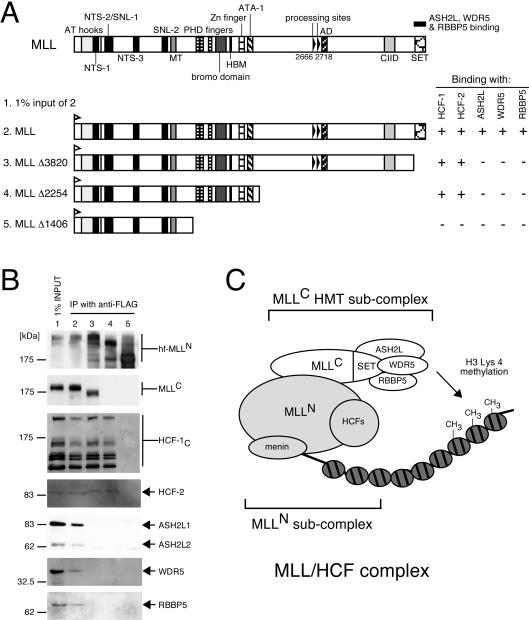
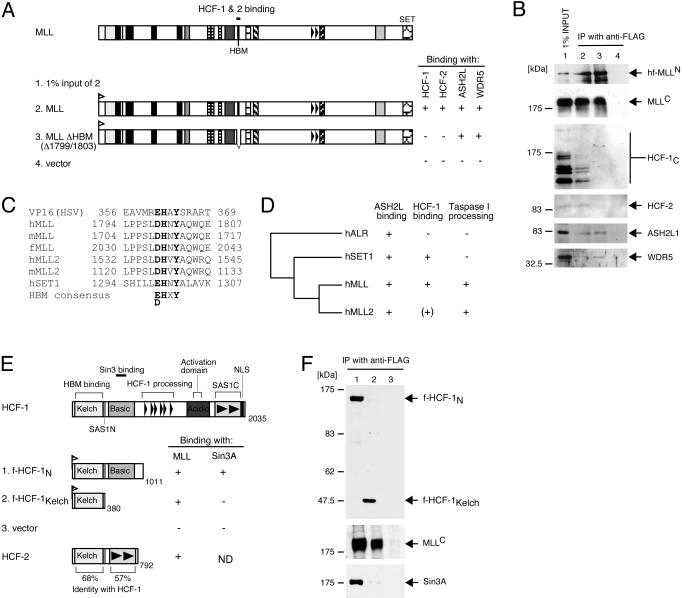
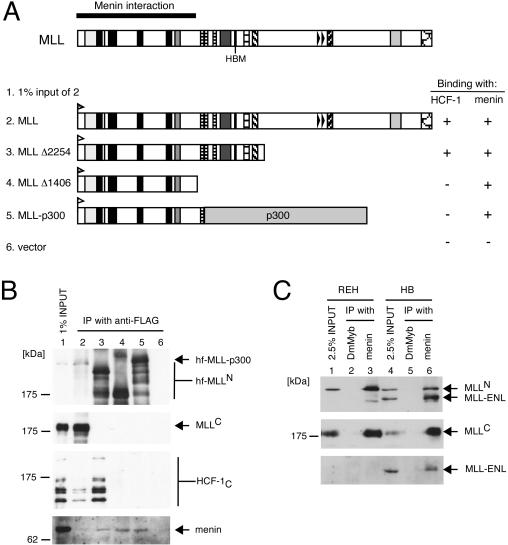
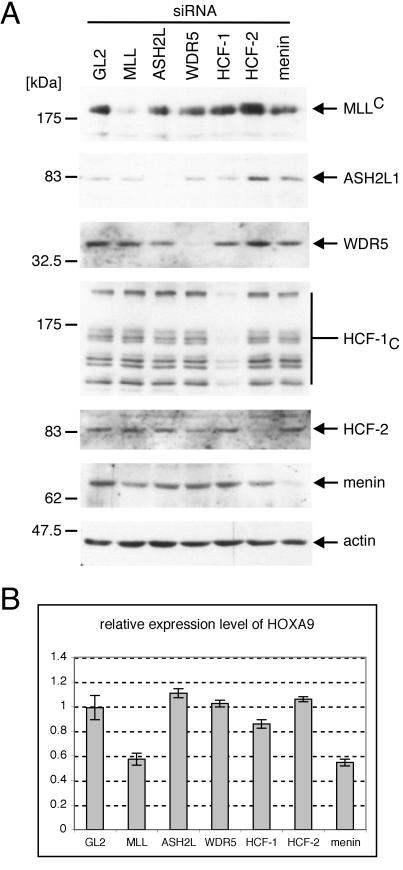
MLL (for mixed-lineage leukemia) is a proto-oncogene that is mutated in a variety of human leukemias. Its product, a homolog of Drosophila melanogaster trithorax, displays intrinsic histone methyltransferase activity and functions genetically to maintain embryonic Hox gene expression. Here we report the biochemical purification of MLL and demonstrate that it associates with a cohort of proteins shared with the yeast and human SET1 histone methyltransferase complexes, including a homolog of Ash2, another Trx-G group protein. Two other members of the novel MLL complex identified here are host cell factor 1 (HCF-1), a transcriptional coregulator, and the related HCF-2, both of which specifically interact with a conserved binding motif in the MLL(N) (p300) subunit of MLL and provide a potential mechanism for regulating its antagonistic transcriptional properties. Menin, a product of the MEN1 tumor suppressor gene, is also a component of the 1-MDa MLL complex. Abrogation of menin expression phenocopies loss of MLL and reveals a critical role for menin in the maintenance of Hox gene expression. Oncogenic mutant forms of MLL retain an ability to interact with menin but not other identified complex components. These studies link the menin tumor suppressor protein with the MLL histone methyltransferase machinery, with implications for Hox gene expression in development and leukemia pathogenesis.
Figures
Similar articles
-
The menin tumor suppressor protein is an essential oncogenic cofactor for MLL-associated leukemogenesis.Cell. 2005 Oct 21;123(2):207-18. doi: 10.1016/j.cell.2005.09.025. Cell. 2005. PMID: 16239140
-
Menin critically links MLL proteins with LEDGF on cancer-associated target genes.Cancer Cell. 2008 Jul 8;14(1):36-46. doi: 10.1016/j.ccr.2008.05.003. Cancer Cell. 2008. PMID: 18598942 Free PMC article.
-
Mechanisms of transformation by MLL.Crit Rev Eukaryot Gene Expr. 2004;14(4):235-54. doi: 10.1615/critreveukaryotgeneexpr.v14.i4.10. Crit Rev Eukaryot Gene Expr. 2004. PMID: 15663355 Review.
-
Menin and MLL cooperatively regulate expression of cyclin-dependent kinase inhibitors.Proc Natl Acad Sci U S A. 2005 Jan 18;102(3):749-54. doi: 10.1073/pnas.0408836102. Epub 2005 Jan 7. Proc Natl Acad Sci U S A. 2005. PMID: 15640349 Free PMC article.
-
MLL: a histone methyltransferase disrupted in leukemia.Trends Mol Med. 2004 Oct;10(10):500-7. doi: 10.1016/j.molmed.2004.08.005. Trends Mol Med. 2004. PMID: 15464450 Review.
Cited by
-
Proteogenomic characterization and mapping of nucleosomes decoded by Brd and HP1 proteins.Genome Biol. 2012 Aug 16;13(8):R68. doi: 10.1186/gb-2012-13-8-r68. Genome Biol. 2012. PMID: 22897906 Free PMC article.
-
A combination of H2A.Z and H4 acetylation recruits Brd2 to chromatin during transcriptional activation.PLoS Genet. 2012;8(11):e1003047. doi: 10.1371/journal.pgen.1003047. Epub 2012 Nov 8. PLoS Genet. 2012. PMID: 23144632 Free PMC article.
-
Assembling a COMPASS.Epigenetics. 2013 Apr;8(4):349-54. doi: 10.4161/epi.24177. Epub 2013 Mar 7. Epigenetics. 2013. PMID: 23470558 Free PMC article.
-
Drosophila melanogaster dHCF interacts with both PcG and TrxG epigenetic regulators.PLoS One. 2011;6(12):e27479. doi: 10.1371/journal.pone.0027479. Epub 2011 Dec 8. PLoS One. 2011. PMID: 22174740 Free PMC article.
-
Regulation by polycomb and trithorax group proteins in Arabidopsis.Arabidopsis Book. 2010;8:e0128. doi: 10.1199/tab.0128. Epub 2010 May 8. Arabidopsis Book. 2010. PMID: 22303254 Free PMC article.
References
-
- Aasland, R., T. J. Gibson, and A. F. Stewart. 1995. The PHD finger: implications for chromatin-mediated transcriptional regulation. Trends Biochem. Sci. 20:56-59. - PubMed
-
- Agarwal, S. K., S. C. Guru, C. Heppner, M. R. Erdos, R. M. Collins, S. Y. Park, S. Saggar, S. C. Chandrasekharappa, F. S. Collins, A. M. Spiegel, S. J. Marx, and A. L. Burns. 1999. Menin interacts with the AP1 transcription factor JunD and represses JunD-activated transcription. Cell 96:143-152. - PubMed
-
- Agarwal, S. K., E. A. Novotny, J. S. Crabtree, J. B. Weitzman, M. Yaniv, A. L. Burns, S. C. Chandrasekharappa, F. S. Collins, A. M. Spiegel, and S. J. Marx. 2003. Transcription factor JunD, deprived of menin, switches from growth suppressor to growth promoter. Proc. Natl. Acad. Sci. USA 100:10770-10775. - PMC - PubMed
-
- Alvarez-Venegas, R., and Z. Avramova. 2002. SET-domain proteins of the Su(var)3-9, E(z) and trithorax families. Gene 285:25-37. - PubMed
-
- Armstrong, S. A., J. E. Staunton, L. B. Silverman, R. Pieters, M. L. den Boer, M. D. Minden, S. E. Sallan, E. S. Lander, T. R. Golub, and S. J. Korsmeyer. 2002. MLL translocations specify a distinct gene expression profile that distinguishes a unique leukemia. Nat. Genet. 30:41-47. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous