

The nuclear scaffold protein NIPP1 is essential for early embryonic development and cell proliferation
- PMID: 15199142
- PMCID: PMC480874
- DOI: 10.1128/MCB.24.13.5863-5874.2004
The nuclear scaffold protein NIPP1 is essential for early embryonic development and cell proliferation
Abstract
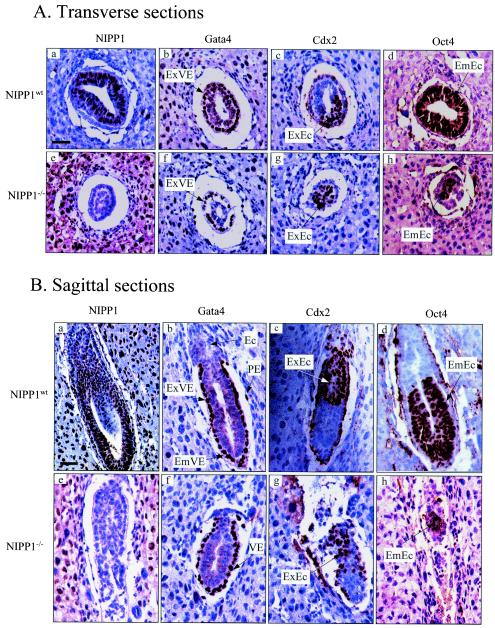
NIPP1 (nuclear inhibitor of protein phosphatase 1) is a ubiquitously expressed nuclear scaffold protein that has been implicated in both transcription and RNA processing. Among its protein ligands are a protein kinase, a protein phosphatase, two splicing factors, and a transcriptional regulator, and the binding of these proteins to NIPP1 is tightly regulated by phosphorylation. To study the function of NIPP1 in vivo, we have used homologous recombination to generate mice that are deficient in NIPP1. NIPP1(-/+) mice developed normally. However, NIPP1(-/-) embryos showed severely retarded growth at embryonic day 6.5 (E6.5) and were resorbed by E8.5. This early embryonic lethality was not associated with increased apoptosis but correlated with impaired cell proliferation. Blastocyst outgrowth experiments and the RNA interference-mediated knockdown of NIPP1 in cultured cells also revealed an essential role for NIPP1 in cell proliferation. In further agreement with this function, no viable NIPP1(-/-) cell lines were obtained by derivation of embryonic stem (ES) cells from blastocysts of NIPP1(-/+) intercrosses or by forced homogenotization of heterozygous ES cells at high concentrations of Geneticin. We conclude that NIPP1 is indispensable for early embryonic development and cell proliferation.
Figures





Similar articles
-
The zinc finger transcription factor 191 is required for early embryonic development and cell proliferation.Exp Cell Res. 2006 Dec 10;312(20):3990-8. doi: 10.1016/j.yexcr.2006.08.020. Epub 2006 Aug 30. Exp Cell Res. 2006. PMID: 17064688
-
Cse1l is essential for early embryonic growth and development.Mol Cell Biol. 2001 Oct;21(20):7020-4. doi: 10.1128/MCB.21.20.7020-7024.2001. Mol Cell Biol. 2001. PMID: 11564884 Free PMC article.
-
mTOR is essential for growth and proliferation in early mouse embryos and embryonic stem cells.Mol Cell Biol. 2004 Aug;24(15):6710-8. doi: 10.1128/MCB.24.15.6710-6718.2004. Mol Cell Biol. 2004. PMID: 15254238 Free PMC article.
-
Polypyrimidine tract-binding protein is essential for early mouse development and embryonic stem cell proliferation.FEBS J. 2009 Nov;276(22):6658-68. doi: 10.1111/j.1742-4658.2009.07380.x. Epub 2009 Oct 16. FEBS J. 2009. PMID: 19843185
-
The protein phosphatase 1 regulator NIPP1 is essential for mammalian spermatogenesis.Sci Rep. 2017 Oct 17;7(1):13364. doi: 10.1038/s41598-017-13809-y. Sci Rep. 2017. PMID: 29042623 Free PMC article.
Cited by
-
Oct4 targets regulatory nodes to modulate stem cell function.PLoS One. 2007 Jun 20;2(6):e553. doi: 10.1371/journal.pone.0000553. PLoS One. 2007. PMID: 17579724 Free PMC article.
-
The NMR structure of the NIPP1 FHA domain.J Biomol NMR. 2008 Mar;40(3):219-24. doi: 10.1007/s10858-008-9222-x. Epub 2008 Feb 6. J Biomol NMR. 2008. PMID: 18253837 No abstract available.
-
The molecular basis for substrate specificity of the nuclear NIPP1:PP1 holoenzyme.Structure. 2012 Oct 10;20(10):1746-56. doi: 10.1016/j.str.2012.08.003. Epub 2012 Aug 30. Structure. 2012. PMID: 22940584 Free PMC article.
-
The split protein phosphatase system.Biochem J. 2018 Dec 6;475(23):3707-3723. doi: 10.1042/BCJ20170726. Biochem J. 2018. PMID: 30523060 Free PMC article. Review.
-
Molecular Signature of HPV-Induced Carcinogenesis: pRb, p53 and Gene Expression Profiling.Curr Genomics. 2009 Mar;10(1):26-34. doi: 10.2174/138920209787581235. Curr Genomics. 2009. PMID: 19721808 Free PMC article.
References
-
- Beullens, M., A. Van Eynde, M. Bollen, and W. Stalmans. 1993. Inactivation of nuclear inhibitory polypeptides of protein phosphatase-1 (NIPP-1) by protein kinase A. J. Biol. Chem. 268:13172-13177. - PubMed
-
- Beullens, M., A. Van Eynde, V. Vulsteke, J. Connor, S. Shenolikar, W. Stalmans, and M. Bollen. 1999. Molecular determinants of nuclear protein phosphatase-1 regulation by NIPP-1. J. Biol. Chem. 274:14053-14061. - PubMed
-
- Beullens, M., A. Van Eynde, W. Stalmans, and M. Bollen. 1992. The isolation of novel inhibitory polypeptides of protein phosphatase 1 from bovine thymus nuclei. J. Biol. Chem. 267:16538-16544. - PubMed
-
- Beullens, M., and M. Bollen. 2002. The protein phosphatase-1 regulator NIPP1 is also a splicing factor involved in a late step of spliceosome assembly. J. Biol. Chem. 277:19855-19860. - PubMed
-
- Beullens, M., V. Vulsteke, A. Van Eynde, I. Jagiello, W. Stalmans, and M. Bollen. 2000. The C-terminus of NIPP1 (nuclear inhibitor of protein phosphatase-1) contains a novel binding site for protein phosphatase-1 that is controlled by tyrosine phosphorylation and RNA binding. Biochem. J. 352:651-658. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases