

Immune functions in mice lacking Clnk, an SLP-76-related adaptor expressed in a subset of immune cells
- PMID: 15199160
- PMCID: PMC480883
- DOI: 10.1128/MCB.24.13.6067-6075.2004
Immune functions in mice lacking Clnk, an SLP-76-related adaptor expressed in a subset of immune cells
Abstract
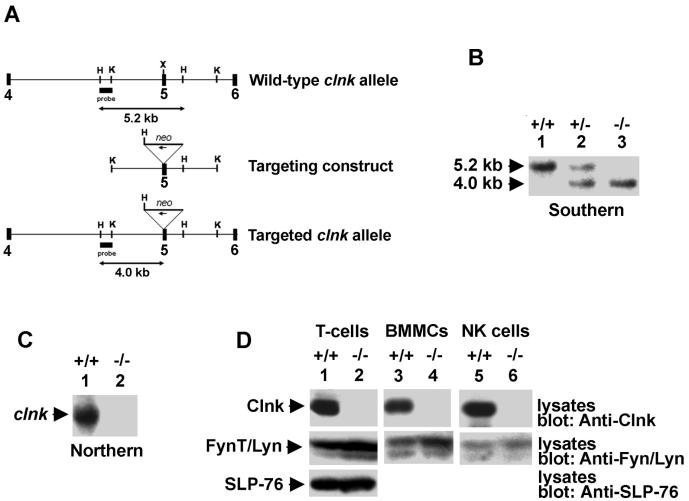
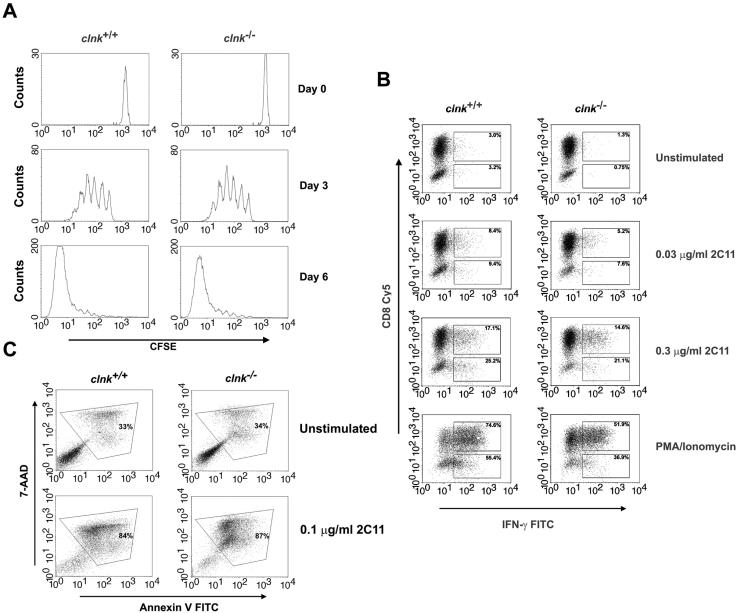
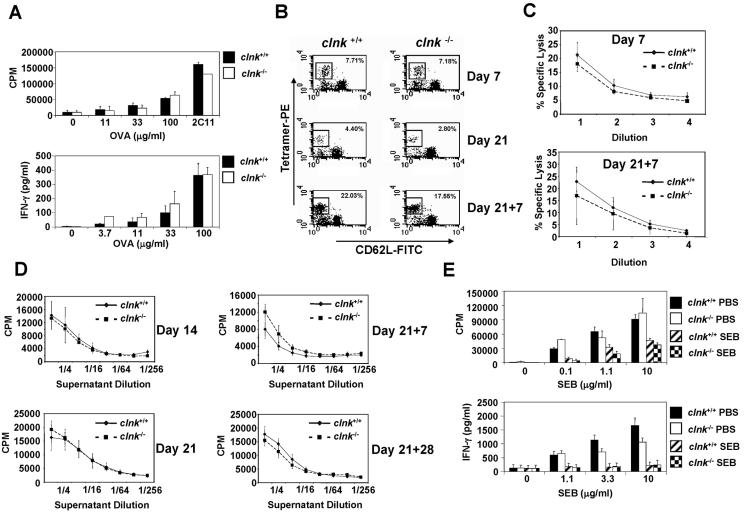
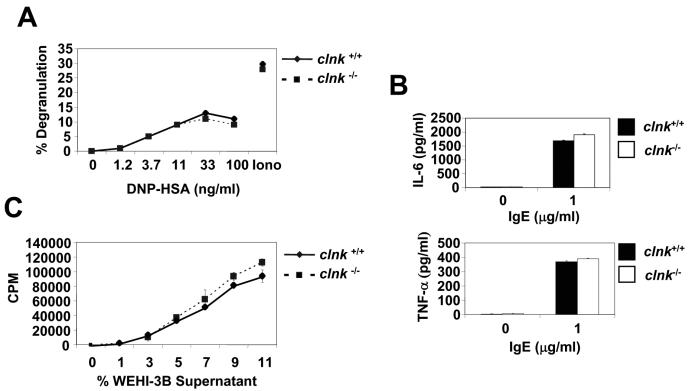
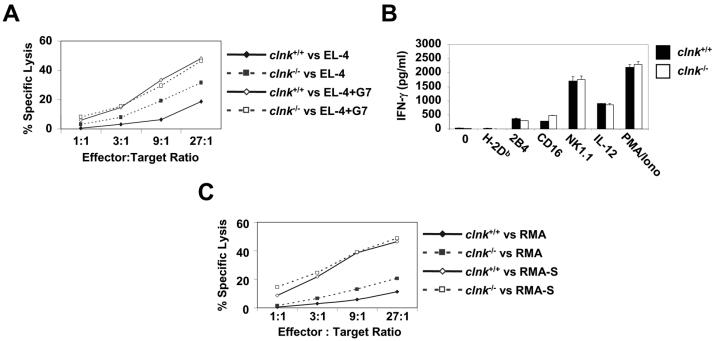
The SLP-76 family of immune cell-specific adaptors is composed of three distinct members named SLP-76, Blnk, and Clnk. They have been implicated in the signaling pathways coupled to immunoreceptors such as the antigen receptors and Fc receptors. Previous studies using gene-targeted mice and deficient cell lines showed that SLP-76 plays a central role in T-cell development and activation. Moreover, it is essential for normal mast cell and platelet activation. In contrast, Blnk is necessary for B-cell development and activation. While the precise function of Clnk is not known, it was reported that Clnk is selectively expressed in mast cells, natural killer (NK) cells, and previously activated T-cells. Moreover, ectopic expression of Clnk was shown to rescue T-cell receptor-mediated signal transduction in an SLP-76-deficient T-cell line, suggesting that, like its relatives, Clnk is involved in the positive regulation of immunoreceptor signaling. Stimulatory effects of Clnk on immunoreceptor signaling were also reported to occur in transfected B-cell and basophil leukemia cell lines. Herein, we attempted to address the physiological role of Clnk in immune cells by the generation of Clnk-deficient mice. The results of our studies demonstrated that Clnk is dispensable for normal differentiation and function of T cells, mast cells, and NK cells. Hence, unlike its relatives, Clnk is not essential for normal immune functions.
Figures
Similar articles
-
BASH-deficient mice: limited primary repertoire and antibody formation, but sufficient affinity maturation and memory B cell generation, in anti-NP response.Int Immunol. 2004 Aug;16(8):1161-71. doi: 10.1093/intimm/dxh116. Epub 2004 Jul 5. Int Immunol. 2004. PMID: 15237108
-
Transcriptional regulation of SLP-76 family hematopoietic cell adaptor MIST/Clnk by STAT5.Biochem Biophys Res Commun. 2004 Aug 13;321(1):145-53. doi: 10.1016/j.bbrc.2004.06.126. Biochem Biophys Res Commun. 2004. PMID: 15358227
-
Distinct regulatory functions of SLP-76 and MIST in NK cell cytotoxicity and IFN-gamma production.Int Immunol. 2008 Mar;20(3):345-52. doi: 10.1093/intimm/dxm150. Epub 2008 Jan 17. Int Immunol. 2008. PMID: 18203684
-
Adaptors as central mediators of signal transduction in immune cells.Nat Immunol. 2003 Feb;4(2):110-6. doi: 10.1038/ni0203-110. Nat Immunol. 2003. PMID: 12555096 Review.
-
Control of lymphocyte development and activation by negative regulatory transmembrane adapter proteins.Immunol Rev. 2008 Aug;224:215-28. doi: 10.1111/j.1600-065X.2008.00656.x. Immunol Rev. 2008. PMID: 18759929 Review.
Cited by
-
Adapters in the organization of mast cell signaling.Immunol Rev. 2009 Nov;232(1):195-217. doi: 10.1111/j.1600-065X.2009.00834.x. Immunol Rev. 2009. PMID: 19909365 Free PMC article. Review.
-
Proteomic analysis of the SH2 domain-containing leukocyte protein of 76 kDa (SLP76) interactome in resting and activated primary mast cells [corrected].Mol Cell Proteomics. 2013 Oct;12(10):2874-89. doi: 10.1074/mcp.M112.025908. Epub 2013 Jul 2. Mol Cell Proteomics. 2013. PMID: 23820730 Free PMC article.
-
The neuroimmune axis of Alzheimer's disease.Genome Med. 2023 Jan 26;15(1):6. doi: 10.1186/s13073-023-01155-w. Genome Med. 2023. PMID: 36703235 Free PMC article. Review.
-
Identification of Immune-Related Gene Signatures in Lung Adenocarcinoma and Lung Squamous Cell Carcinoma.Front Immunol. 2021 Nov 23;12:752643. doi: 10.3389/fimmu.2021.752643. eCollection 2021. Front Immunol. 2021. PMID: 34887858 Free PMC article.
-
IFN-γ induction by neutrophil-derived IL-17A homodimer augments pulmonary antibacterial defense.Mucosal Immunol. 2016 May;9(3):718-29. doi: 10.1038/mi.2015.95. Epub 2015 Sep 9. Mucosal Immunol. 2016. PMID: 26349661 Free PMC article.
References
-
- Abtahian, F., A. Guerriero, E. Sebzda, M. M. Lu, R. Zhou, A. Mocsai, E. E. Myers, B. Huang, D. G. Jackson, V. A. Ferrari, V. Tybulewicz, C. A. Lowell, J. J. Lepore, G. A. Koretzky, and M. L. Kahn. 2003. Regulation of blood and lymphatic vascular separation by signaling proteins SLP-76 and Syk. Science 299:247-251. - PMC - PubMed
-
- Bertram, E. M., P. Lau, and T. H. Watts. 2002. Temporal segregation of 4-1BB versus CD28-mediated costimulation: 4-1BB ligand influences T cell numbers late in the primary response and regulates the size of the T cell memory response following influenza infection. J. Immunol. 168:3777-3785. - PubMed
-
- Biassoni, R., C. Cantoni, D. Pende, S. Sivori, S. Parolini, M. Vitale, C. Bottino, and A. Moretta. 2001. Human natural killer cell receptors and co-receptors. Immunol. Rev. 181:203-214. - PubMed
-
- Brandle, D., K. Burki, V. A. Wallace, U. H. Rohrer, T. W. Mak, B. Malissen, H. Hengartner, and H. Pircher. 1991. Involvement of both T cell receptor V alpha and V beta variable region domains and alpha chain junctional region in viral antigen recognition. Eur. J. Immunol. 21:2195-2202. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases