

The single-strand DNA binding activity of human PC4 prevents mutagenesis and killing by oxidative DNA damage
- PMID: 15199162
- PMCID: PMC480877
- DOI: 10.1128/MCB.24.13.6084-6093.2004
The single-strand DNA binding activity of human PC4 prevents mutagenesis and killing by oxidative DNA damage
Abstract
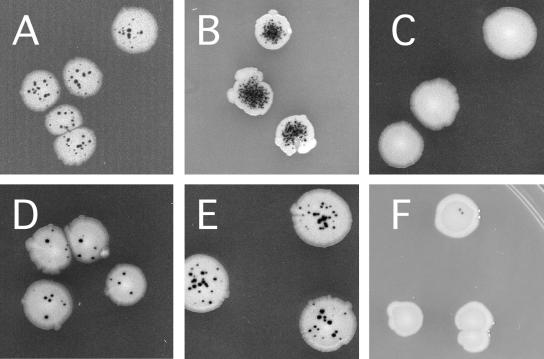
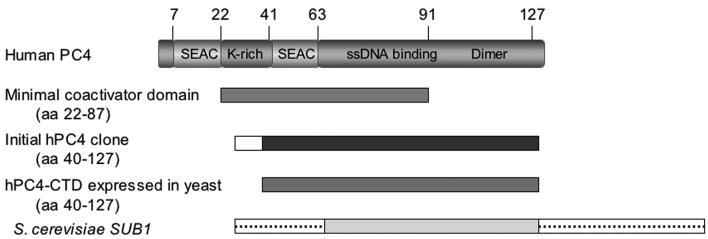
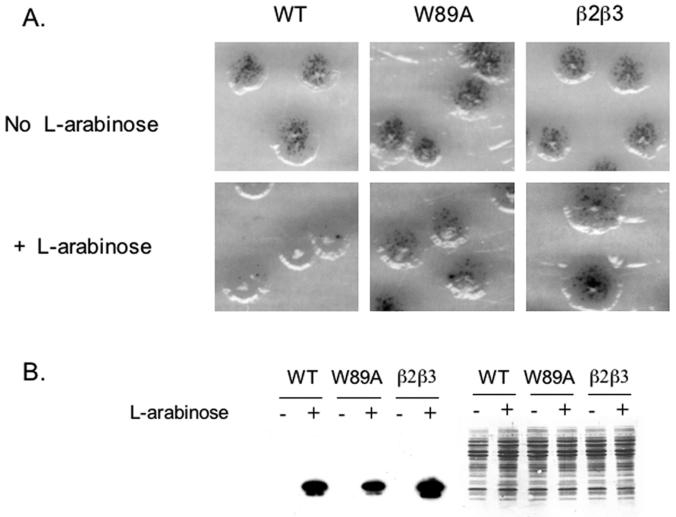
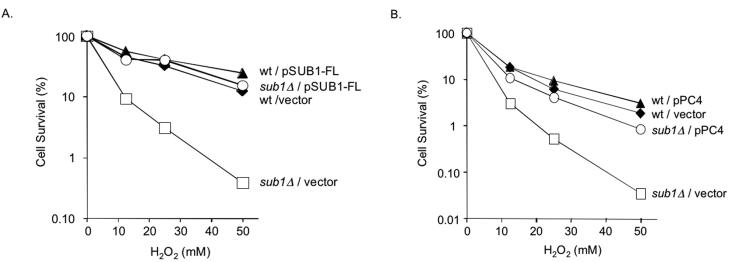
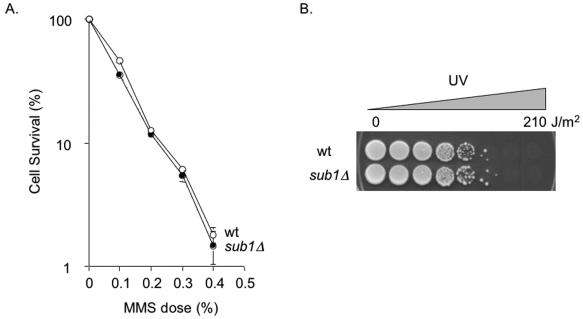
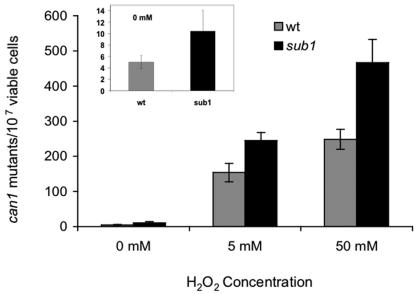
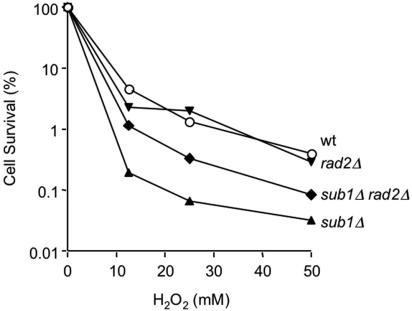
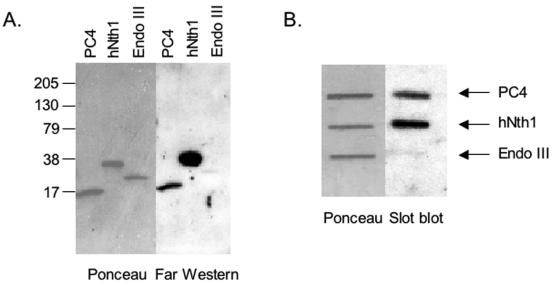
Human positive cofactor 4 (PC4) is a transcriptional coactivator with a highly conserved single-strand DNA (ssDNA) binding domain of unknown function. We identified PC4 as a suppressor of the oxidative mutator phenotype of the Escherichia coli fpg mutY mutant and demonstrate that this suppression requires its ssDNA binding activity. Saccharomyces cerevisiae mutants lacking their PC4 ortholog Sub1 are sensitive to hydrogen peroxide and exhibit spontaneous and peroxide-induced hypermutability. PC4 expression suppresses the peroxide sensitivity of the yeast sub1Delta mutant, suggesting that the human protein has a similar function. A role for yeast and human proteins in DNA repair is suggested by the demonstration that Sub1 acts in a peroxide resistance pathway involving Rad2 and by the physical interaction of PC4 with the human Rad2 homolog XPG. We show that XPG recruits PC4 to a bubble-containing DNA substrate with a resulting displacement of XPG and formation of a PC4-DNA complex. We discuss the possible requirement for PC4 in either global or transcription-coupled repair of oxidative DNA damage to mediate the release of XPG bound to its substrate.
Figures
Similar articles
-
Functional characterization of the interacting domains of the positive coactivator PC4 with the transcription factor AP-2alpha.Gene. 2003 Nov 27;320:155-64. doi: 10.1016/s0378-1119(03)00823-0. Gene. 2003. PMID: 14597399
-
Yeast SUB1 is a suppressor of TFIIB mutations and has homology to the human co-activator PC4.EMBO J. 1996 Apr 15;15(8):1933-40. EMBO J. 1996. PMID: 8617240 Free PMC article.
-
High-affinity DNA binding by the C-terminal domain of the transcriptional coactivator PC4 requires simultaneous interaction with two opposing unpaired strands and results in helix destabilization.J Mol Biol. 1998 Feb 20;276(2):367-77. doi: 10.1006/jmbi.1997.1534. J Mol Biol. 1998. PMID: 9512709
-
Sub1/PC4 a chromatin associated protein with multiple functions in transcription.RNA Biol. 2010 May-Jun;7(3):287-90. doi: 10.4161/rna.7.3.11491. Epub 2010 May 9. RNA Biol. 2010. PMID: 20305379 Review.
-
Sub1/PC4, a multifaceted factor: from transcription to genome stability.Curr Genet. 2017 Dec;63(6):1023-1035. doi: 10.1007/s00294-017-0715-6. Epub 2017 May 31. Curr Genet. 2017. PMID: 28567479 Review.
Cited by
-
Effect of labeling with iron oxide particles or nanodiamonds on the functionality of adipose-derived mesenchymal stem cells.PLoS One. 2013;8(1):e52997. doi: 10.1371/journal.pone.0052997. Epub 2013 Jan 3. PLoS One. 2013. PMID: 23301012 Free PMC article.
-
The transcriptional coactivator PC4/Sub1 has multiple functions in RNA polymerase II transcription.EMBO J. 2005 Mar 9;24(5):1009-20. doi: 10.1038/sj.emboj.7600575. Epub 2005 Feb 3. EMBO J. 2005. PMID: 15692559 Free PMC article.
-
Genome-Wide Association Study Reveals PC4 as the Candidate Gene for Thermal Tolerance in Bay Scallop (Argopecten irradians irradians).Front Genet. 2021 Jul 19;12:650045. doi: 10.3389/fgene.2021.650045. eCollection 2021. Front Genet. 2021. PMID: 34349776 Free PMC article.
-
SUB1 Plays a Negative Role during Starvation Induced Sporulation Program in Saccharomyces cerevisiae.PLoS One. 2015 Jul 6;10(7):e0132350. doi: 10.1371/journal.pone.0132350. eCollection 2015. PLoS One. 2015. PMID: 26147804 Free PMC article.
-
Inhibition of PC4 radiosensitizes non-small cell lung cancer by transcriptionally suppressing XLF.Cancer Med. 2018 Apr;7(4):1326-1337. doi: 10.1002/cam4.1332. Epub 2018 Mar 9. Cancer Med. 2018. PMID: 29522271 Free PMC article.
References
-
- Aburatani, H., Y. Hippo, T. Ishida, R. Takashima, C. Matsuba, T. Kodama, M. Takano, A. Yasui, K. Yamamoto, M. Asano, K. Fukasawa, T. Yoshinari, H. Inoue, E. Ohtsuka, and S. Nisimura. 1997. Cloning and characterization of mammalian 8-hydroyguanine-specific DNA glycosylase/apurinic, apyrimidinic lyase, a functional mutM homologue. Cancer Res. 57:2151-2156. - PubMed
-
- Arai, K., K. Morishita, K. Shinmura, T. Kohno, S. R. Kim, T. Nohmi, M. Taniwaki, S. Ohwada, and J. Yokota. 1997. Cloning of a human homolog of the yeast OGG1 gene that is involved in the repair of oxidative DNA damage. Oncogene 14:2857-2861. - PubMed
-
- Bohm, M., H. Kirch, T. Otto, H. Rubben, and I. Wieland. 1997. Deletion analysis at the DEL-27, APC and MTS1 loci in bladder cancer: LOH at the DEL-27 locus on 5p13-12 is a prognostic marker of tumor progression. Int. J. Cancer 74:291-295. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous