

Invertebrate immune systems--not homogeneous, not simple, not well understood
- PMID: 15199951
- PMCID: PMC5426807
- DOI: 10.1111/j.0105-2896.2004.0117.x
Invertebrate immune systems--not homogeneous, not simple, not well understood
Abstract
The approximate 30 extant invertebrate phyla have diversified along separate evolutionary trajectories for hundreds of millions of years. Although recent work understandably has emphasized the commonalities of innate defenses, there is also ample evidence, as from completed genome studies, to suggest that even members of the same invertebrate order have taken significantly different approaches to internal defense. These data suggest that novel immune capabilities will be found among the different phyla. Many invertebrates have intimate associations with symbionts that may play more of a role in internal defense than generally appreciated. Some invertebrates that are either long lived or have colonial body plans may diversify components of their defense systems via somatic mutation. Somatic diversification following pathogen exposure, as seen in plants, has been investigated little in invertebrates. Recent molecular studies of sponges, cnidarians, shrimp, mollusks, sea urchins, tunicates, and lancelets have found surprisingly diversified immune molecules, and a model is presented that supports the adaptive value of diversified non-self recognition molecules in invertebrates. Interactions between invertebrates and viruses also remain poorly understood. As we are in the midst of alarming losses of coral reefs, increased pathogen challenge to invertebrate aquaculture, and rampant invertebrate-transmitted parasites of humans and domestic animals, we need a better understanding of invertebrate immunology.
Figures
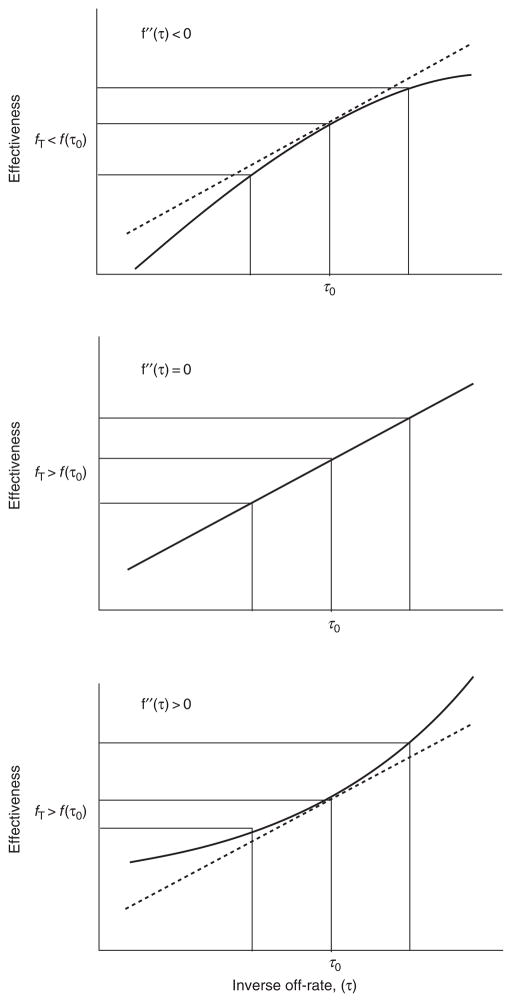
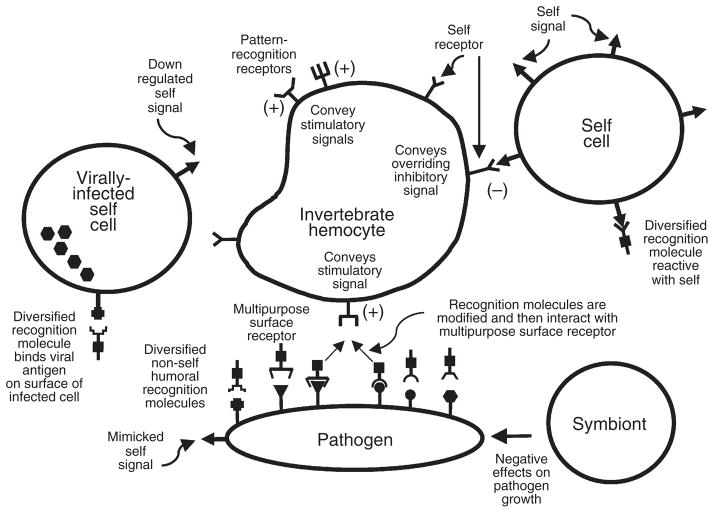
References
-
- Hoffmann JA, Kafatos FC, Janeway CA, Ezekowitz RAB. Phylogenetic perspectives in innate immunity. Science. 1999;284:1313–1318. - PubMed
-
- Hoffmann JA, Reichhart JM. Drosophila innate immunity: an evolutionary perspective. Nat Immunol. 2002;3:121–126. - PubMed
-
- Medzhitov R, Preston-Hurlburt P, Janeway CA. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature. 1997;388:394–397. - PubMed
-
- Takeda K, Akira S. Toll receptors and pathogen resistance. Cell Microbiol. 2003;5:143–153. - PubMed
-
- Schnare M, Barton GM, Holt AC, Takeda K, Akira S, Medzhitov R. Toll-like receptors control activation of adaptive immune responses. Nat Immunol. 2001;2:947–950. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
