

The YopD translocator of Yersinia pseudotuberculosis is a multifunctional protein comprised of discrete domains
- PMID: 15205412
- PMCID: PMC421591
- DOI: 10.1128/JB.186.13.4110-4123.2004
The YopD translocator of Yersinia pseudotuberculosis is a multifunctional protein comprised of discrete domains
Abstract
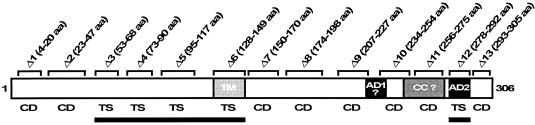
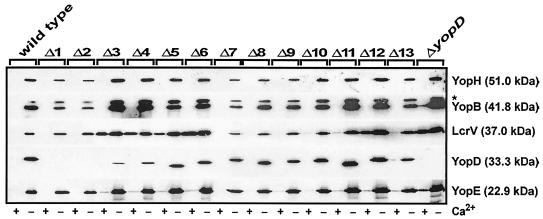
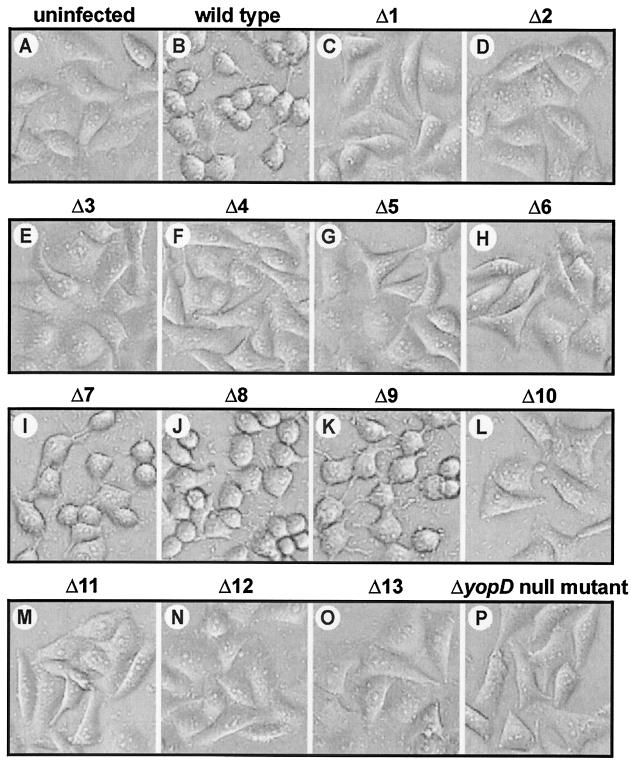
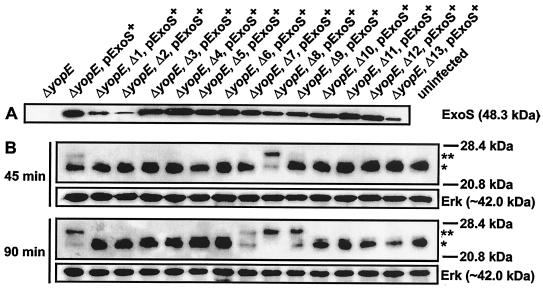
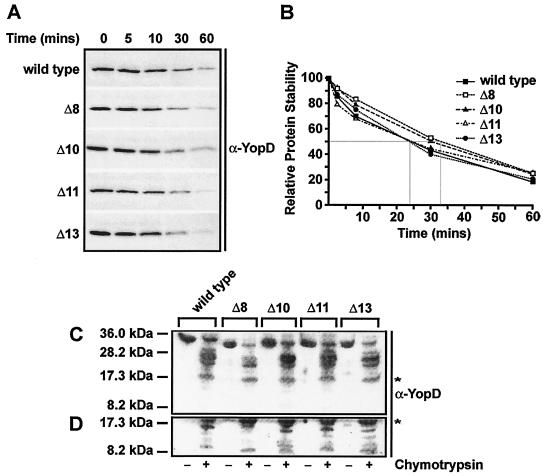
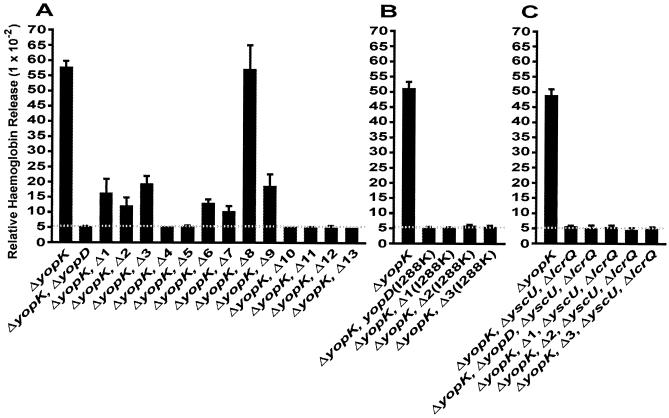
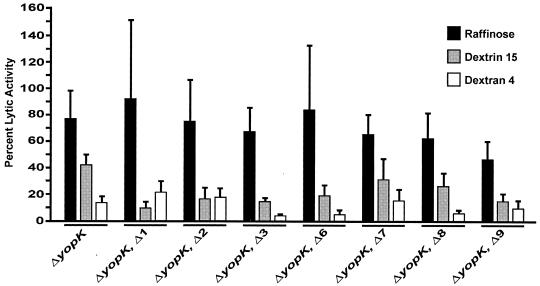
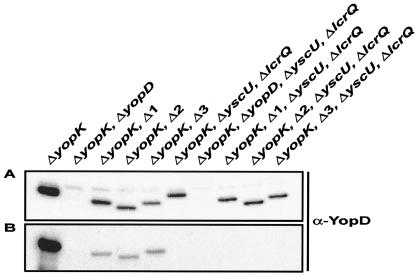
To establish an infection, Yersinia pseudotuberculosis utilizes a plasmid-encoded type III translocon to microinject several anti-host Yop effectors into the cytosol of target eukaryotic cells. YopD has been implicated in several key steps during Yop effector translocation, including maintenance of yop regulatory control and pore formation in the target cell membrane through which effectors traverse. These functions are mediated, in part, by an interaction with the cognate chaperone, LcrH. To gain insight into the complex molecular mechanisms of YopD function, we performed a systematic mutagenesis study to search for discrete functional domains. We highlighted amino acids beyond the first three N-terminal residues that are dispensable for YopD secretion and confirmed that an interaction between YopD and LcrH is essential for maintenance of yop regulatory control. In addition, discrete domains within YopD that are essential for both pore formation and translocation of Yop effectors were identified. Significantly, other domains were found to be important for effector microinjection but not for pore formation. Therefore, YopD is clearly essential for several discrete steps during efficient Yop effector translocation. Recognition of this modular YopD domain structure provides important insights into the function of YopD.
Figures
Similar articles
-
Minimal YopB and YopD translocator secretion by Yersinia is sufficient for Yop-effector delivery into target cells.Microbes Infect. 2007 Feb;9(2):224-33. doi: 10.1016/j.micinf.2006.11.010. Epub 2006 Dec 18. Microbes Infect. 2007. PMID: 17223369
-
The type III secretion chaperone LcrH co-operates with YopD to establish a negative, regulatory loop for control of Yop synthesis in Yersinia pseudotuberculosis.Mol Microbiol. 2001 Nov;42(4):1075-93. doi: 10.1046/j.1365-2958.2001.02702.x. Mol Microbiol. 2001. PMID: 11737648
-
Coiled-coils in the YopD translocator family: a predicted structure unique to the YopD N-terminus contributes to full virulence of Yersinia pseudotuberculosis.Infect Genet Evol. 2012 Dec;12(8):1729-42. doi: 10.1016/j.meegid.2012.07.016. Epub 2012 Aug 11. Infect Genet Evol. 2012. PMID: 22910185
-
The Yersinia Yop virulon, a bacterial system to subvert cells of the primary host defense.Folia Microbiol (Praha). 1998;43(3):253-61. doi: 10.1007/BF02818610. Folia Microbiol (Praha). 1998. PMID: 9717252 Review.
-
The Yersinia Yop virulon: a bacterial system for subverting eukaryotic cells.Mol Microbiol. 1997 Mar;23(5):861-7. doi: 10.1046/j.1365-2958.1997.2731623.x. Mol Microbiol. 1997. PMID: 9076724 Review.
Cited by
-
LcrV mutants that abolish Yersinia type III injectisome function.J Bacteriol. 2013 Feb;195(4):777-87. doi: 10.1128/JB.02021-12. Epub 2012 Dec 7. J Bacteriol. 2013. PMID: 23222719 Free PMC article.
-
A bacterial secreted translocator hijacks riboregulators to control type III secretion in response to host cell contact.PLoS Pathog. 2019 Jun 7;15(6):e1007813. doi: 10.1371/journal.ppat.1007813. eCollection 2019 Jun. PLoS Pathog. 2019. PMID: 31173606 Free PMC article.
-
Yersinia controls type III effector delivery into host cells by modulating Rho activity.PLoS Pathog. 2008 Jan;4(1):e3. doi: 10.1371/journal.ppat.0040003. PLoS Pathog. 2008. PMID: 18193942 Free PMC article.
-
Inflammasome activation in response to the Yersinia type III secretion system requires hyperinjection of translocon proteins YopB and YopD.mBio. 2015 Feb 17;6(1):e02095-14. doi: 10.1128/mBio.02095-14. mBio. 2015. PMID: 25691590 Free PMC article.
-
Random mutagenesis identifies a C-terminal region of YopD important for Yersinia type III secretion function.PLoS One. 2015 Mar 25;10(3):e0120471. doi: 10.1371/journal.pone.0120471. eCollection 2015. PLoS One. 2015. PMID: 25807250 Free PMC article.
References
-
- Aepfelbacher, M., and J. Heesemann. 2001. Modulation of Rho GTPases and the actin cytoskeleton by Yersinia outer proteins (Yops). Int. J. Med. Microbiol. 291:269-276. - PubMed
-
- Anderson, D. M., and O. Schneewind. 1999. Type III machines of Gram-negative pathogens: injecting virulence factors into host cells and more. Curr. Opin. Microbiol. 2:18-24. - PubMed
-
- Barbieri, J. T. 2000. Pseudomonas aeruginosa exoenzyme S, a bifunctional type-III secreted cytotoxin. Int. J. Med. Microbiol. 290:381-387. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
