

Telomere exchange between linear replicons of Borrelia burgdorferi
- PMID: 15205414
- PMCID: PMC421586
- DOI: 10.1128/JB.186.13.4134-4141.2004
Telomere exchange between linear replicons of Borrelia burgdorferi
Abstract
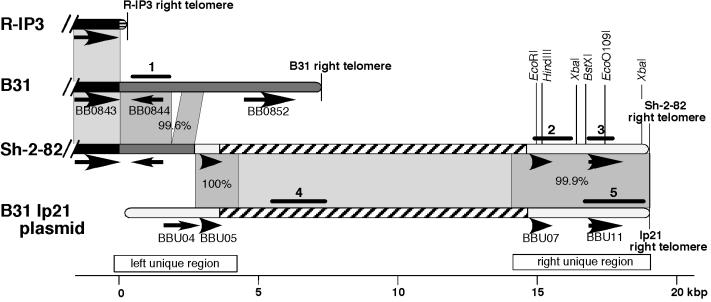
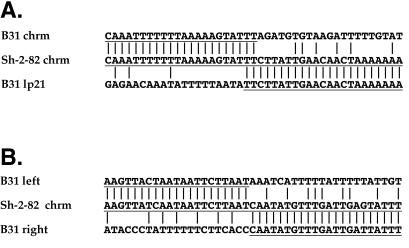
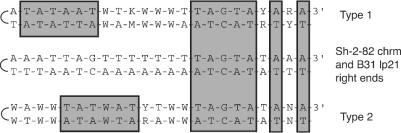
Spirochetes in the genus Borrelia carry a linear chromosome and numerous linear plasmids that have covalently closed hairpin telomeres. The overall organization of the large chromosome of Borrelia burgdorferi appears to have been quite stable over recent evolutionary time; however, a large fraction of natural isolates carry differing lengths of DNA that extend the right end of the chromosome between about 7 and 20 kbp relative to the shortest chromosomes. We present evidence here that a rather recent nonhomologous recombination event in the B. burgdorferi strain Sh-2-82 lineage has replaced its right chromosomal telomere with a large portion of the linear plasmid lp21, which is present in the strain B31 lineage. At least two successive rounds of addition of linear plasmid genetic material to the chromosomal right end appear to have occurred at the Sh-2-82 right telomere, suggesting that this is an evolutionary mechanism by which plasmid genetic material can become part of the chromosome. The unusual nonhomologous nature of this rearrangement suggests that, barring horizontal transfer, it can be used as a unique genetic marker for this lineage of B. burgdorferi chromosomes.
Figures
Similar articles
-
Hairpin telomeres and genome plasticity in Borrelia: all mixed up in the end.Mol Microbiol. 2005 Nov;58(3):625-35. doi: 10.1111/j.1365-2958.2005.04872.x. Mol Microbiol. 2005. PMID: 16238614 Review.
-
Evolution of the linear DNA replicons of the Borrelia spirochetes.Curr Opin Microbiol. 1999 Oct;2(5):529-34. doi: 10.1016/s1369-5274(99)00012-0. Curr Opin Microbiol. 1999. PMID: 10508719 Review.
-
Plasmid diversity and phylogenetic consistency in the Lyme disease agent Borrelia burgdorferi.BMC Genomics. 2017 Feb 15;18(1):165. doi: 10.1186/s12864-017-3553-5. BMC Genomics. 2017. PMID: 28201991 Free PMC article.
-
Linear plasmids of Borrelia burgdorferi have a telomeric structure and sequence similar to those of a eukaryotic virus.J Bacteriol. 1991 Nov;173(22):7233-9. doi: 10.1128/jb.173.22.7233-7239.1991. J Bacteriol. 1991. PMID: 1938918 Free PMC article.
-
Telomeres of the linear chromosomes of Lyme disease spirochaetes: nucleotide sequence and possible exchange with linear plasmid telomeres.Mol Microbiol. 1997 Nov;26(3):581-96. doi: 10.1046/j.1365-2958.1997.6051963.x. Mol Microbiol. 1997. PMID: 9402027
Cited by
-
Comparative genomics of Streptomyces avermitilis, Streptomyces cattleya, Streptomyces maritimus and Kitasatospora aureofaciens using a Streptomyces coelicolor microarray system.Antonie Van Leeuwenhoek. 2008 Jan-Feb;93(1-2):1-25. doi: 10.1007/s10482-007-9175-1. Epub 2007 Jun 21. Antonie Van Leeuwenhoek. 2008. PMID: 17588127 Free PMC article.
-
Characterization and genomic analysis of the Lyme disease spirochete bacteriophage ϕBB-1.bioRxiv [Preprint]. 2024 Jan 13:2024.01.08.574763. doi: 10.1101/2024.01.08.574763. bioRxiv. 2024. Update in: PLoS Pathog. 2024 Apr 1;20(4):e1012122. doi: 10.1371/journal.ppat.1012122. PMID: 38260690 Free PMC article. Updated. Preprint.
-
Differential telomere processing by Borrelia telomere resolvases in vitro but not in vivo.J Bacteriol. 2006 Nov;188(21):7378-86. doi: 10.1128/JB.00760-06. Epub 2006 Aug 25. J Bacteriol. 2006. PMID: 16936037 Free PMC article.
-
Whole genome sequencing of Borrelia miyamotoi isolate Izh-4: reference for a complex bacterial genome.BMC Genomics. 2020 Jan 6;21(1):16. doi: 10.1186/s12864-019-6388-4. BMC Genomics. 2020. PMID: 31906865 Free PMC article.
-
Genome stability of Lyme disease spirochetes: comparative genomics of Borrelia burgdorferi plasmids.PLoS One. 2012;7(3):e33280. doi: 10.1371/journal.pone.0033280. Epub 2012 Mar 14. PLoS One. 2012. PMID: 22432010 Free PMC article.
References
-
- Busch, U., C. Hizo-Teufel, R. Bohmer, V. Fingerle, D. Rossler, B. Wilske, and V. Preac-Mursic. 1996. Borrelia burgdorferi sensu lato strains isolated from cutaneous Lyme borreliosis biopsies differentiated by pulsed-field gel electrophoresis. Scand. J. Infect. Dis. 28:583-589. - PubMed
-
- Casjens, S. 1998. The diverse and dynamic structure of bacterial genomes. Annu. Rev. Genet. 32:339-377. - PubMed
-
- Casjens, S. 1999. Evolution of the linear DNA replicons of the Borrelia spirochetes. Curr. Opin. Microbiol. 2:529-534. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
