

CBP histone acetyltransferase activity is a critical component of memory consolidation
- PMID: 15207240
- PMCID: PMC8048715
- DOI: 10.1016/j.neuron.2004.06.002
CBP histone acetyltransferase activity is a critical component of memory consolidation
Abstract
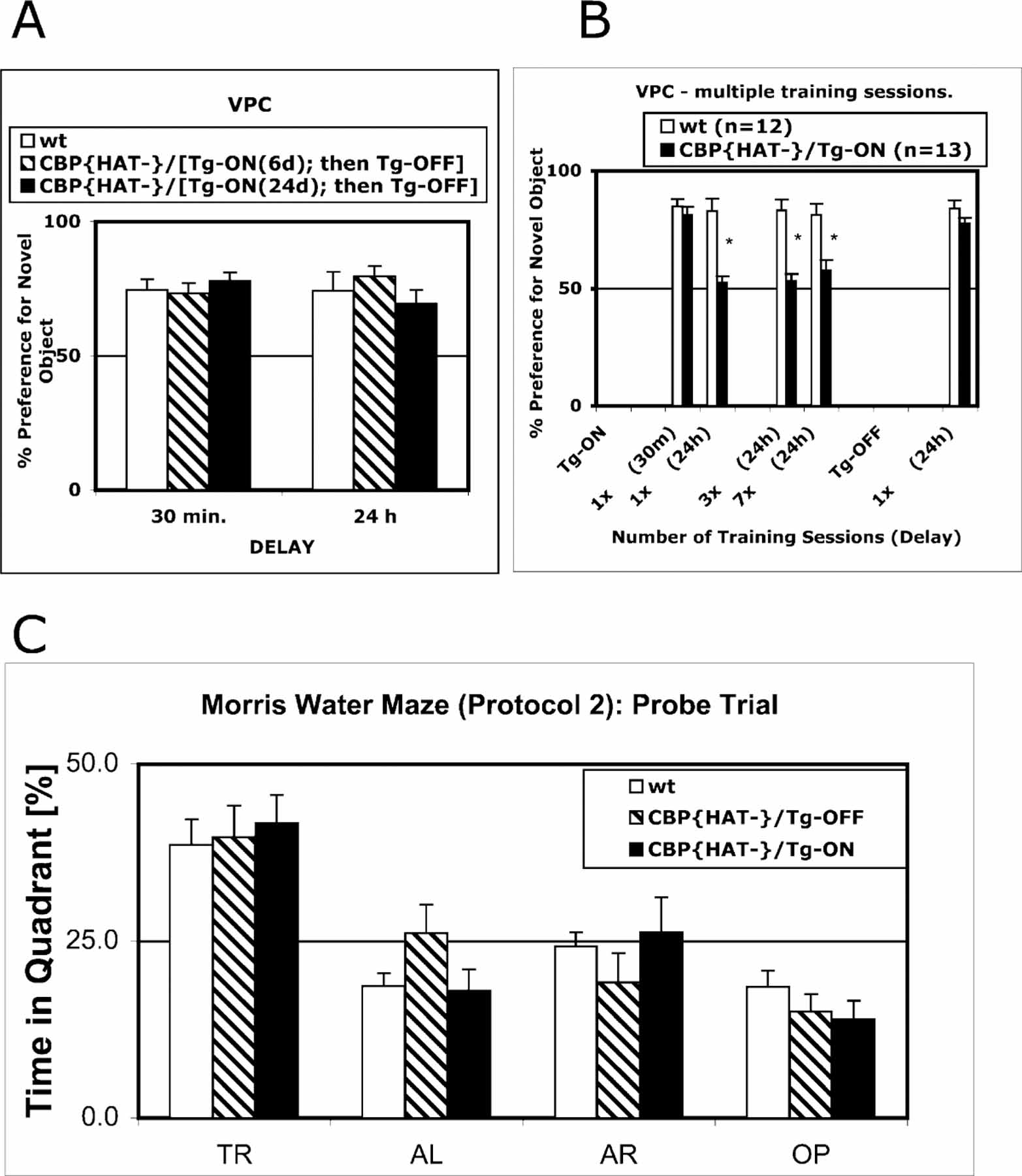
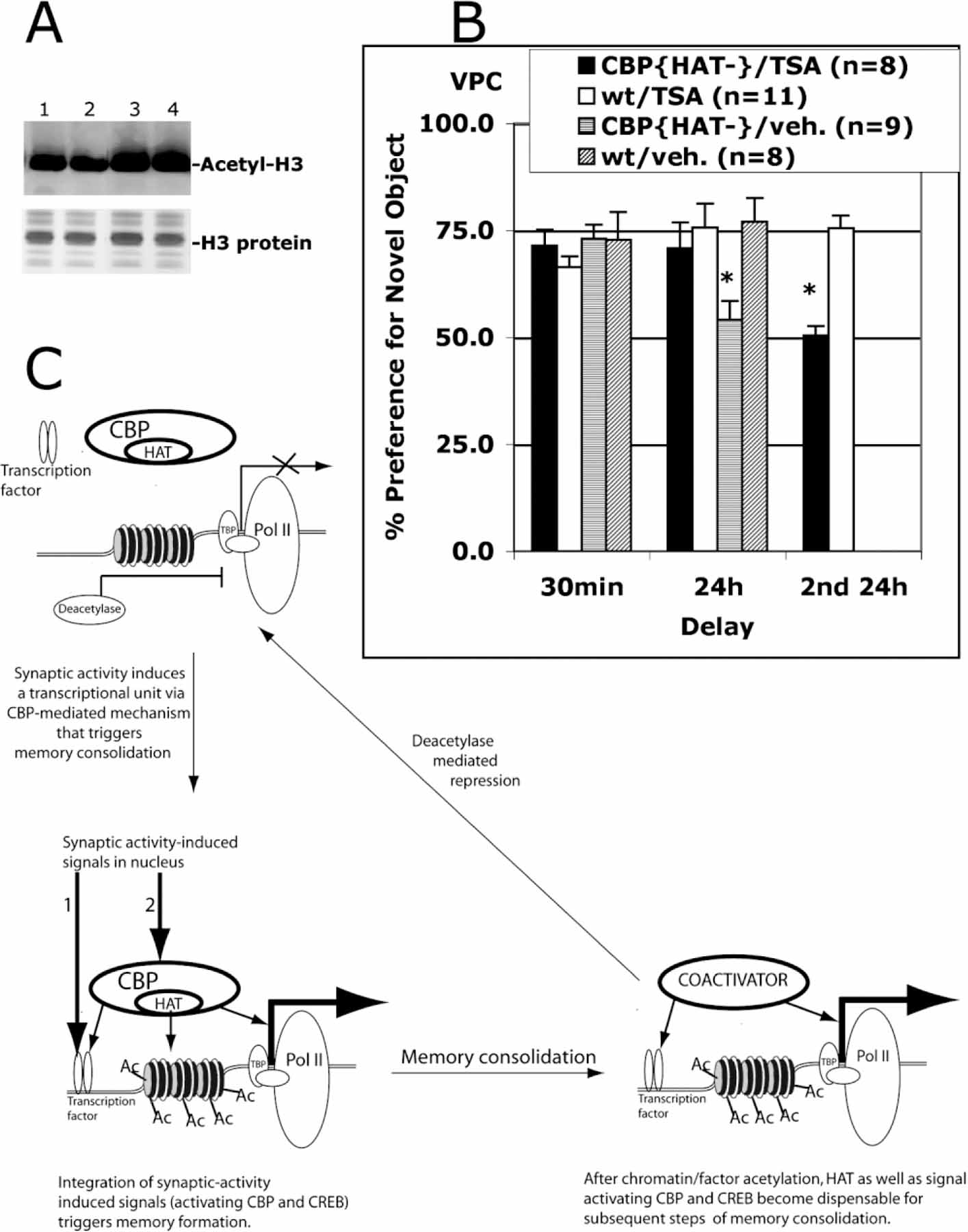
The stabilization of learned information into long-term memories requires new gene expression. CREB binding protein (CBP) is a coactivator of transcription that can be independently regulated in neurons. CBP functions both as a platform for recruiting other required components of the transcriptional machinery and as a histone acetyltransferase (HAT) that alters chromatin structure. To dissect the chromatin remodeling versus platform function of CBP or the developmental versus adult role of this gene, we generated transgenic mice that express CBP in which HAT activity is eliminated. Acquisition of new information and short-term memory is spared in these mice, while the stabilization of short-term memory into long-term memory is impaired. The behavioral phenotype is due to an acute requirement for CBP HAT activity in the adult as it is rescued by both suppression of transgene expression or by administration of the histone deacetylase inhibitor Trichostatin A (TSA) in adult animals.
Copyright 2004 Cell Press
Figures
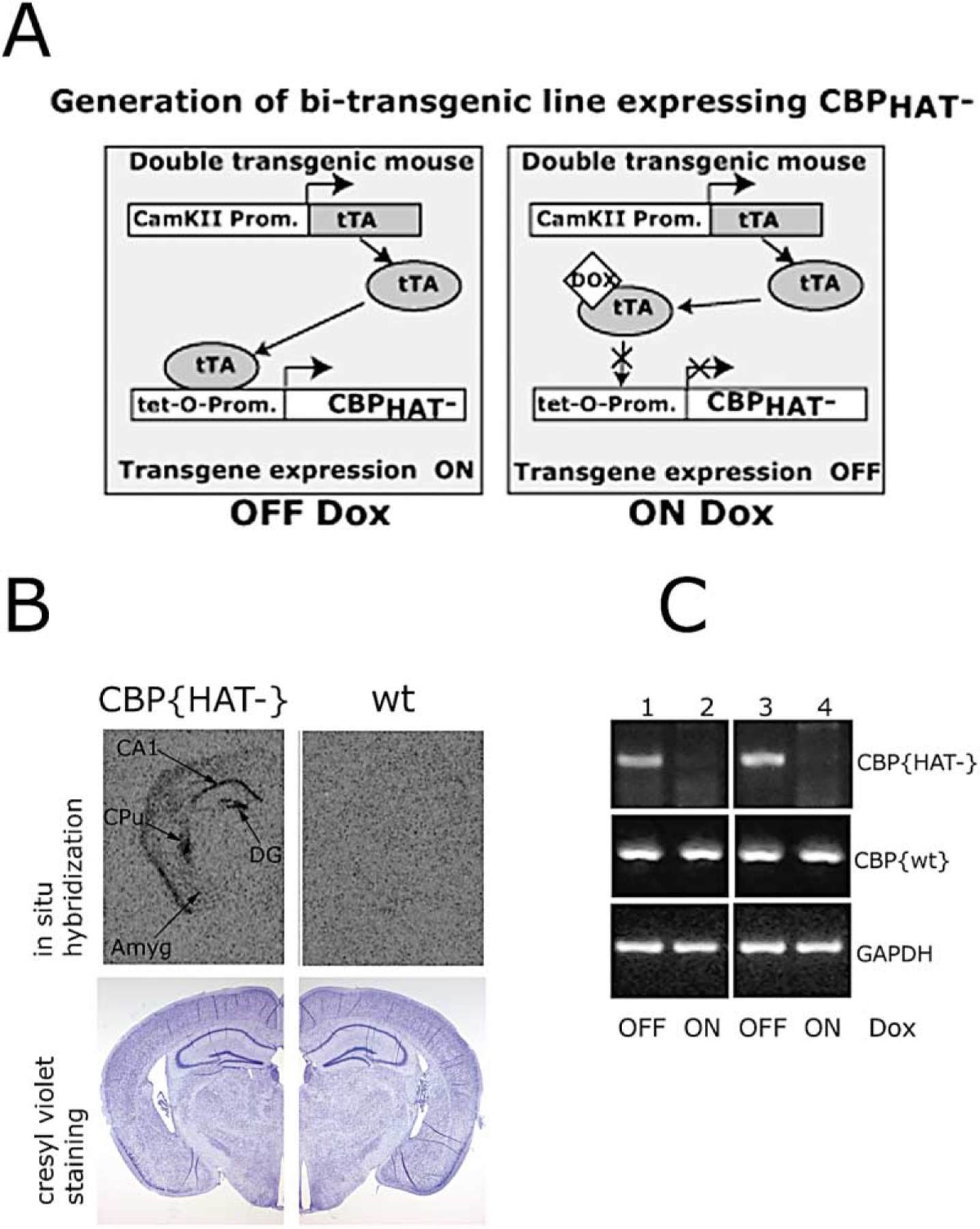
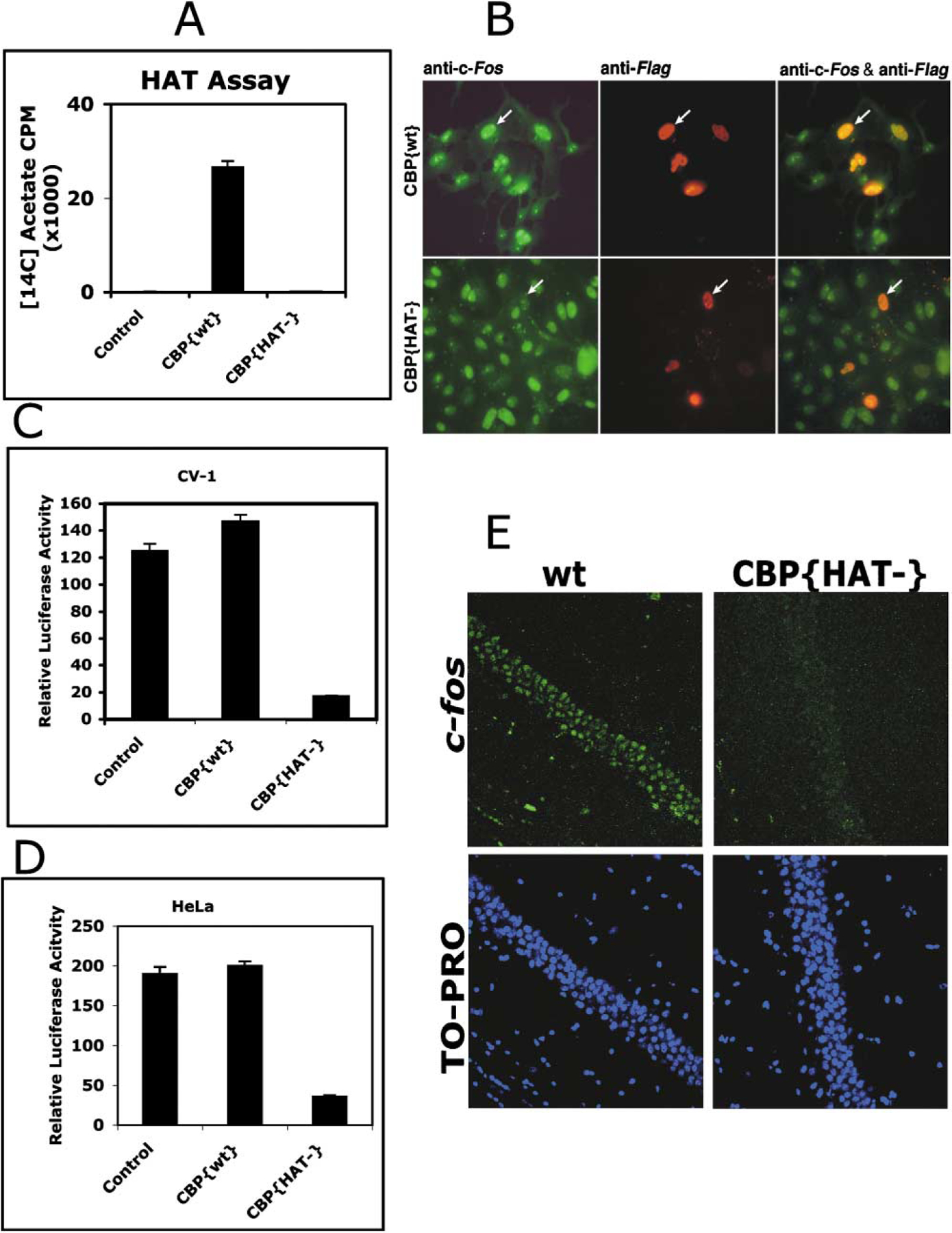
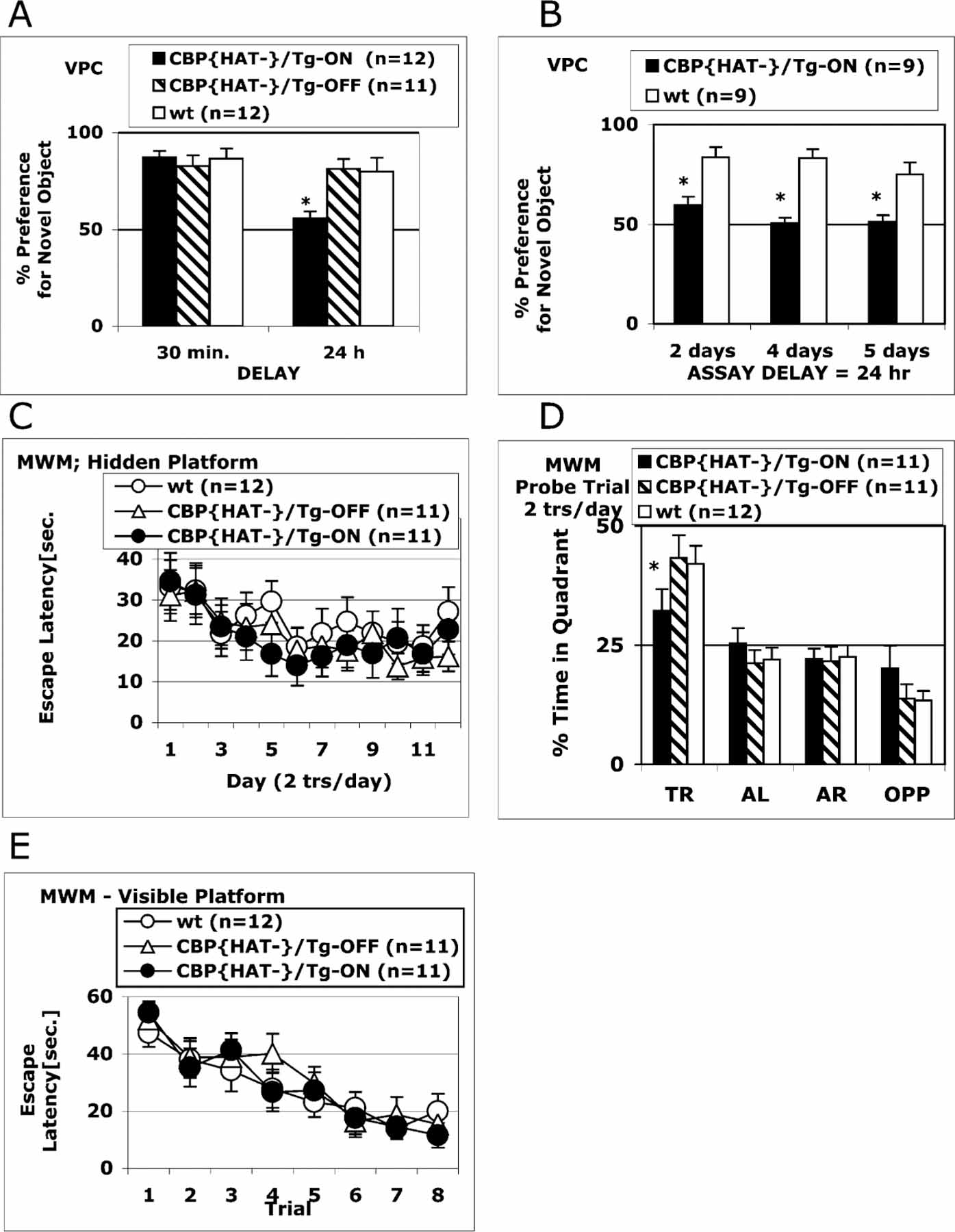
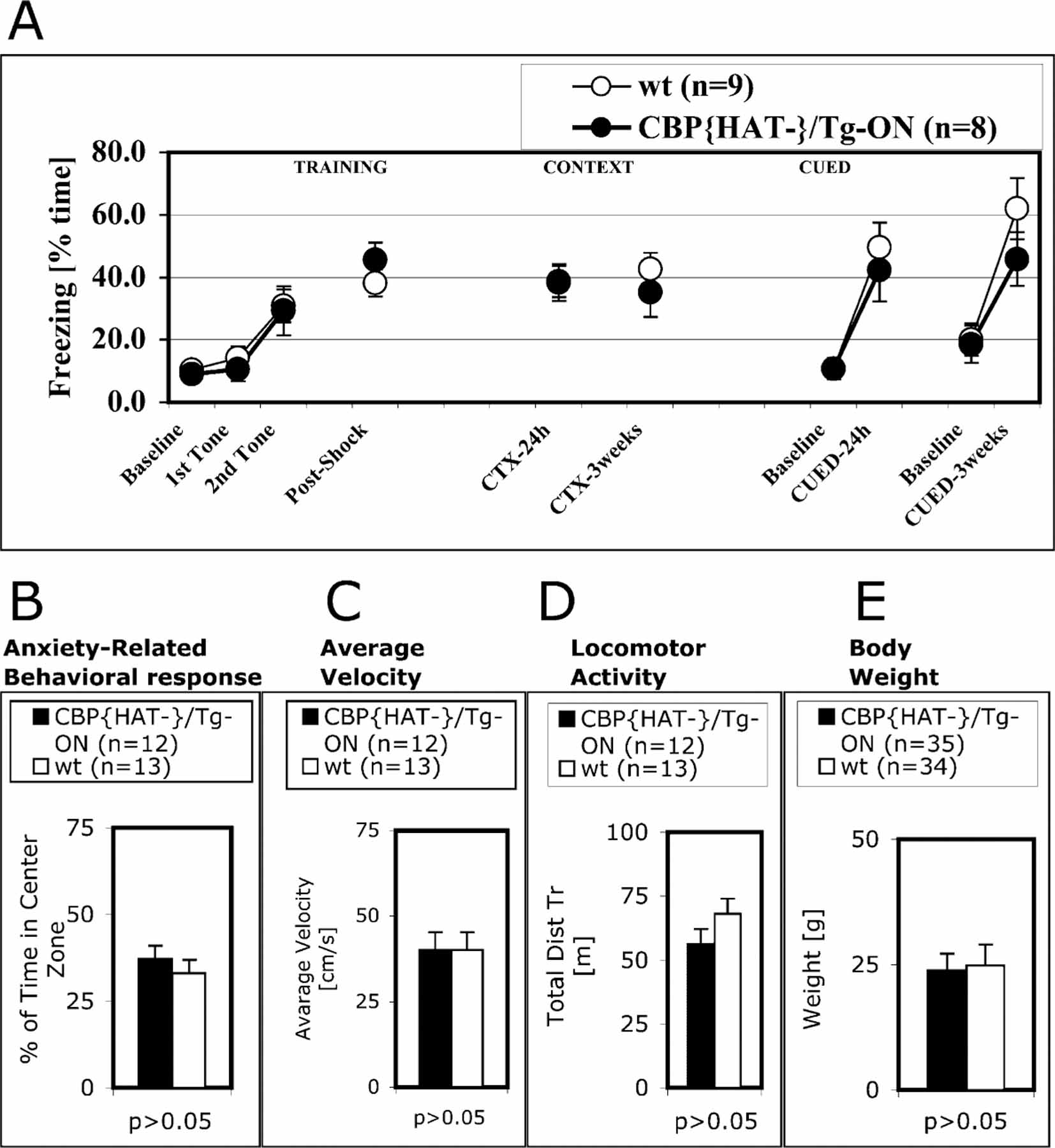
Comment in
-
To learn better, keep the HAT on.Neuron. 2004 Jun 24;42(6):879-81. doi: 10.1016/j.neuron.2004.06.007. Neuron. 2004. PMID: 15207231
Similar articles
-
Chromatin acetylation, memory, and LTP are impaired in CBP+/- mice: a model for the cognitive deficit in Rubinstein-Taybi syndrome and its amelioration.Neuron. 2004 Jun 24;42(6):947-59. doi: 10.1016/j.neuron.2004.05.021. Neuron. 2004. PMID: 15207239
-
To learn better, keep the HAT on.Neuron. 2004 Jun 24;42(6):879-81. doi: 10.1016/j.neuron.2004.06.007. Neuron. 2004. PMID: 15207231
-
Long-term memory deficits in Huntington's disease are associated with reduced CBP histone acetylase activity.Hum Mol Genet. 2012 Mar 15;21(6):1203-16. doi: 10.1093/hmg/ddr552. Epub 2011 Nov 24. Hum Mol Genet. 2012. PMID: 22116937
-
CBP and p300: HATs for different occasions.Biochem Pharmacol. 2004 Sep 15;68(6):1145-55. doi: 10.1016/j.bcp.2004.03.045. Biochem Pharmacol. 2004. PMID: 15313412 Review.
-
The versatile functions of the transcriptional coactivators p300 and CBP and their roles in disease.Histol Histopathol. 2002 Apr;17(2):657-68. doi: 10.14670/HH-17.657. Histol Histopathol. 2002. PMID: 11962765 Review.
Cited by
-
Acetyl-CoA Metabolism and Histone Acetylation in the Regulation of Aging and Lifespan.Antioxidants (Basel). 2021 Apr 8;10(4):572. doi: 10.3390/antiox10040572. Antioxidants (Basel). 2021. PMID: 33917812 Free PMC article. Review.
-
The potential of epigenetics in stress-enhanced fear learning models of PTSD.Learn Mem. 2016 Sep 15;23(10):576-86. doi: 10.1101/lm.040485.115. Print 2016 Oct. Learn Mem. 2016. PMID: 27634148 Free PMC article. Review.
-
From Linkage Studies to Epigenetics: What We Know and What We Need to Know in the Neurobiology of Schizophrenia.Front Neurosci. 2016 May 11;10:202. doi: 10.3389/fnins.2016.00202. eCollection 2016. Front Neurosci. 2016. PMID: 27242407 Free PMC article. Review.
-
Transcriptome Analysis of iPSC-Derived Neurons from Rubinstein-Taybi Patients Reveals Deficits in Neuronal Differentiation.Mol Neurobiol. 2020 Sep;57(9):3685-3701. doi: 10.1007/s12035-020-01983-6. Epub 2020 Jun 20. Mol Neurobiol. 2020. PMID: 32562237 Free PMC article.
-
Molecular Genetic Strategies in the Study of Corticohippocampal Circuits.Cold Spring Harb Perspect Biol. 2015 Jul 1;7(7):a021725. doi: 10.1101/cshperspect.a021725. Cold Spring Harb Perspect Biol. 2015. PMID: 26134320 Free PMC article. Review.
References
-
- Andrew RJ (1980). The functional organization of phases of memory consolidation. In Advances in the Study of Behaviour, Hinde RA, Beer C, and Bunsel M, eds. (New York: Academic Press; ), pp. 337–367.
-
- Bachevalier J (1990). Ontogenetic development of habit and memory formation in primates. Ann. N Y Acad. Sci 608, 457–484. - PubMed
-
- Bading H, Ginty DD, and Greenberg ME (1993). Regulation of gene expression in hippocampal neurons by distinct calcium signaling pathways. Science 260, 181–186. - PubMed
-
- Bito H, Deisseroth K, and Tsien RW (1996). CREB phosphorylation and dephosphorylation: a Ca(2+)- and stimulus duration-dependent switch for hippocampal gene expression. Cell 87, 1203–1214. - PubMed
-
- Bourtchuladze R, Frenguelli B, Blendy J, Cioffi D, Schutz G, and Silva AJ (1994). Deficient long-term memory in mice with a targeted mutation of the cAMP-responsive element-binding protein. Cell 79, 59–68. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
