

Arabidopsis ETA2, an apparent ortholog of the human cullin-interacting protein CAND1, is required for auxin responses mediated by the SCF(TIR1) ubiquitin ligase
- PMID: 15208392
- PMCID: PMC514168
- DOI: 10.1105/tpc.021923
Arabidopsis ETA2, an apparent ortholog of the human cullin-interacting protein CAND1, is required for auxin responses mediated by the SCF(TIR1) ubiquitin ligase
Abstract
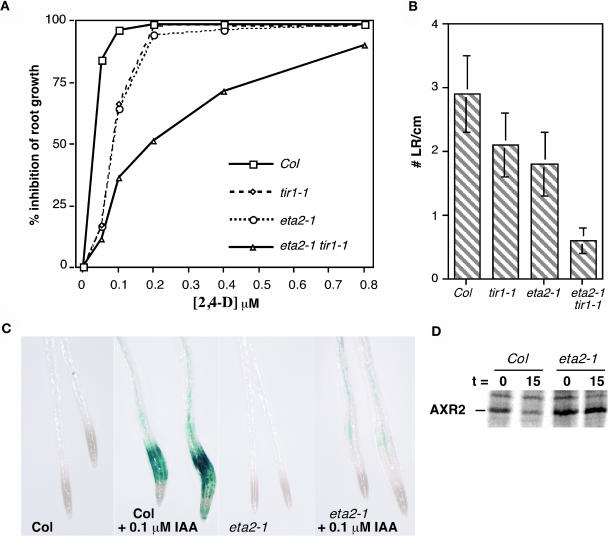
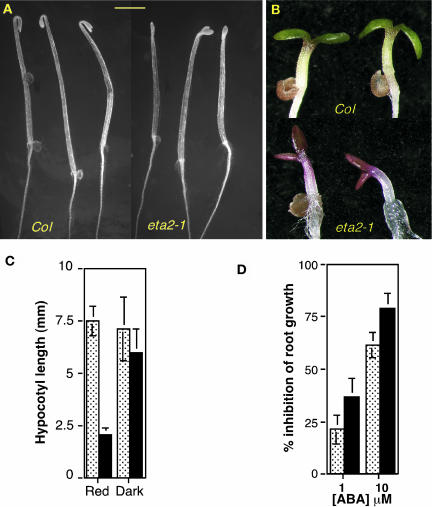
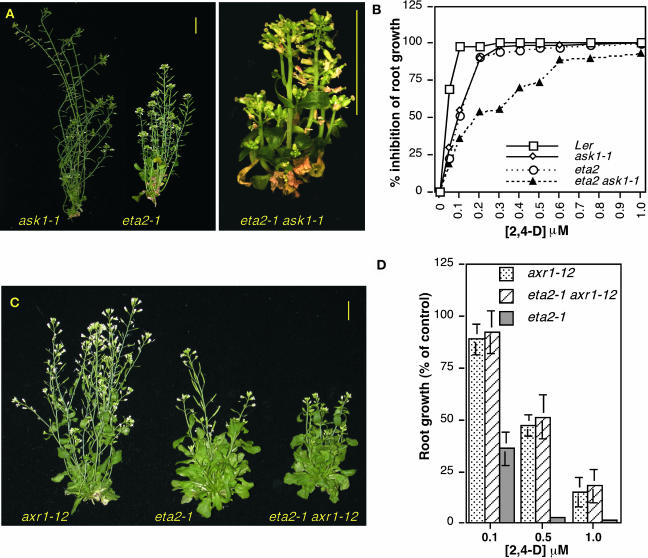
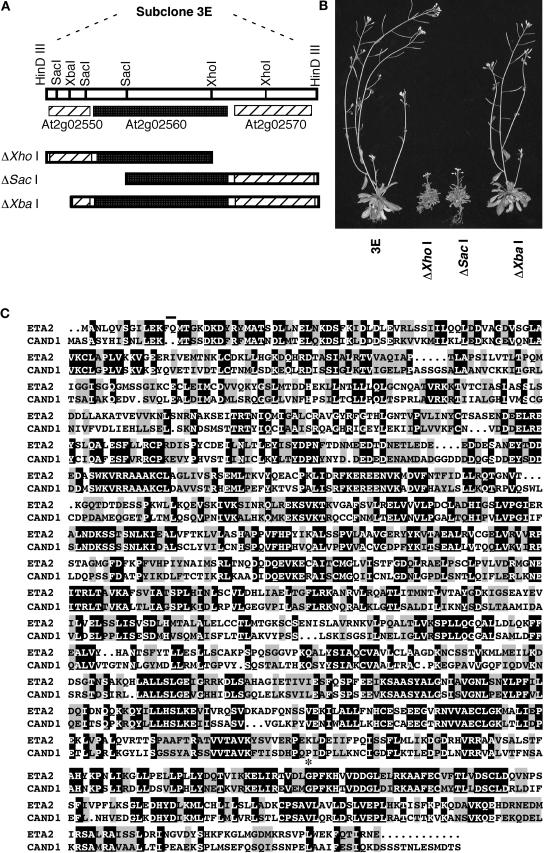
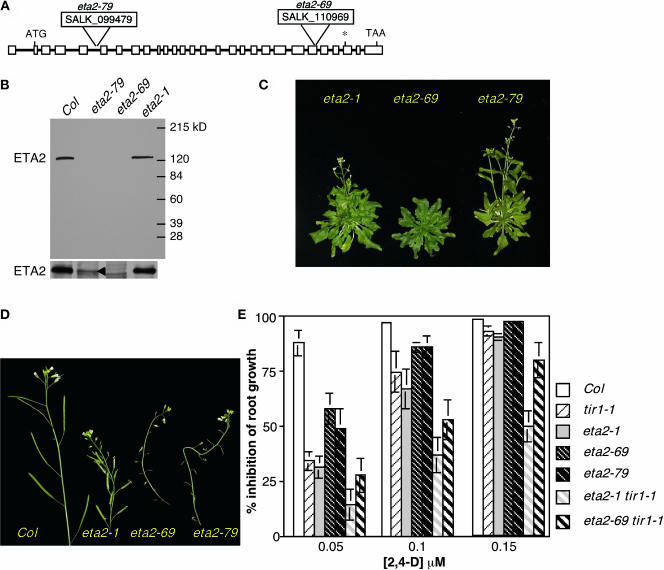
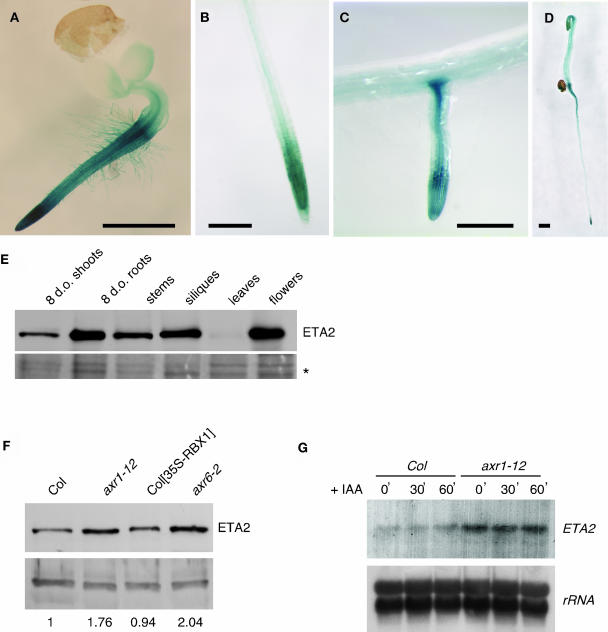
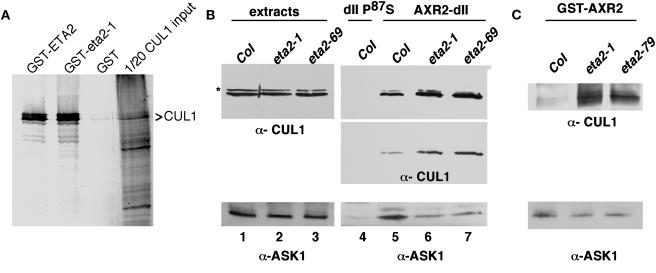
Auxin response in Arabidopsis thaliana requires the SCF(TIR1) ubiquitin ligase. In response to the hormone, SCF(TIR1) targets members of the auxin/indoleacetic acid (Aux/IAA) family of transcriptional regulators for ubiquitin-mediated proteolysis. To identify additional regulators of SCF(TIR1) activity, we conducted a genetic screen to isolate enhancers of the tir1-1 auxin response defect. Here, we report our analysis of the eta2 mutant. Mutations in ETA2 confer several phenotypes consistent with reduced auxin response. ETA2 encodes the Arabidopsis ortholog of human Cullin Associated and Neddylation-Dissociated (CAND1)/TIP120A, a protein recently identified as a cullin-interacting factor. Previous biochemical studies of CAND1 have suggested that it specifically binds to unmodified CUL1 to negatively regulate SCF assembly. By contrast, we find that ETA2 positively regulates SCF(TIR1) because Aux/IAA protein stability is significantly increased in eta2 mutants. Modification of CUL1 by the RUB1/NEDD8 ubiquitin-like protein has been proposed to free CUL1 from CAND1 and promote SCF assembly. We present double mutant analyses of eta2 axr1 plants indicating that liberating CUL1 from ETA2/CAND1 is not the primary role of the RUB modification pathway in the regulation of SCF activity. Our genetic and molecular analysis of SCF(TIR1) function in eta2 mutants provides novel insight into the role of CAND1 in the regulation of SCF ubiquitin-ligase activity.
Figures

References
-
- Alonso, J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301, 653–657. - PubMed
-
- Cope, G.A., and Deshaies, R.J. (2003). COP9 signalosome: A multifunctional regulator of SCF and other cullin-based ubiquitin ligases. Cell 114, 663–671. - PubMed
-
- Cope, G.A., Suh, G.S., Aravind, L., Schwarz, S.E., Zipursky, S.L., Koonin, E.V., and Deshaies, R.J. (2002). Role of predicted metalloprotease motif of Jab1/Csn5 in cleavage of Nedd8 from Cul1. Science 298, 608–611. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
