

Review
doi: 10.1104/pp.104.042614.
The molecular basis of diversity in the photoperiodic flowering responses of Arabidopsis and rice
Affiliations
- PMID: 15208414
- PMCID: PMC514104
- DOI: 10.1104/pp.104.042614
Item in Clipboard
Review
The molecular basis of diversity in the photoperiodic flowering responses of Arabidopsis and rice
Plant Physiol.
2004 Jun.
No abstract available
Figures
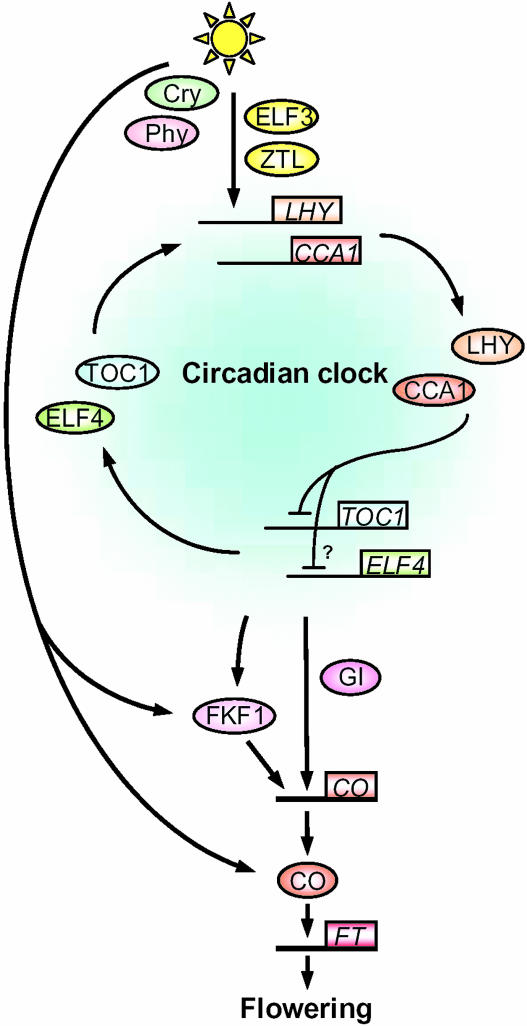
Model of the circadian system of Arabidopsis and its relationship to the flowering-time gene CO. Phytochromes and cryptochromes perceive light and are involved in resetting of the circadian clock. ELF3 and ZTL mediate between photoreceptors and the circadian clock. LHY/CCA1 and TOC1/ELF4 form a negative feedback loop within the circadian oscillator. LHY/CCA1 act as negative regulators of TOC1 and ELF4, which positively regulate the transcription of LHY/CCA1. The oscillator functions to determine the phase of CO transcription, a key gene that mediates between the circadian clock and flowering. The transcription of CO is regulated by FKF1 and GI, whose transcription is under the control of the circadian clock. FKF1 protein is directly regulated by light, and this allows FKF1 to increase CO transcript under LDs. CO protein is also directly activated by light, and this allows CO to generate a LD signal and activate a flowering-time gene FT for the promotion of flowering specifically under LDs.
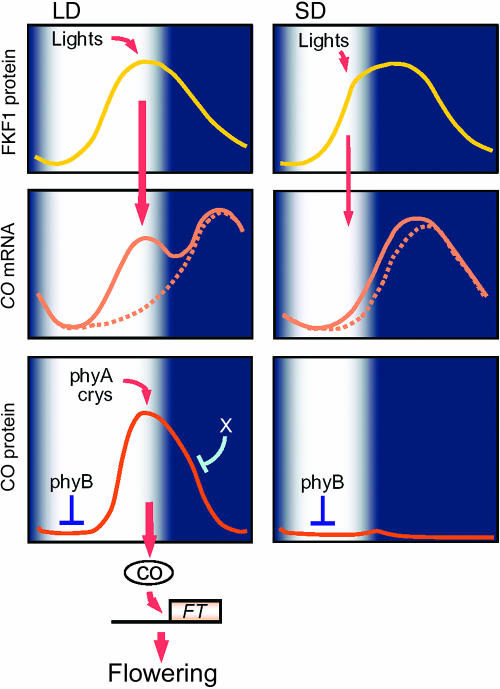
A model of daylength measurement in Arabidopsis. Expression of FKF1 protein is regulated by the circadian clock and exhibits a diurnal rhythm under LD and SD. FKF1 protein functions as a photoreceptor, and accumulates at high levels during mid to end of the day under an LD. FKF1 is regulated by light and functions under LDs to increase CO mRNA abundance. CO mRNA levels in an fkf1 mutant are indicated by the dotted line, whereas the solid line illustrates CO mRNA levels in wild-type plants. CO protein is thereafter activated by light, because blue and far-red light stabilize CO through the action of cryptochromes and phyA and darkness destabilizes it. phyB antagonizes the activity of phyA and cryptochromes and promotes the degradation of CO especially in the morning, allowing CO protein to be expressed with a more refined waveform under LDs. The combination of phyB activity that promotes degradation of CO in the morning and FKF1 activity raising CO mRNA levels during the day under LDs results in robust FT induction and floral promotion specifically under LDs in Arabidopsis.
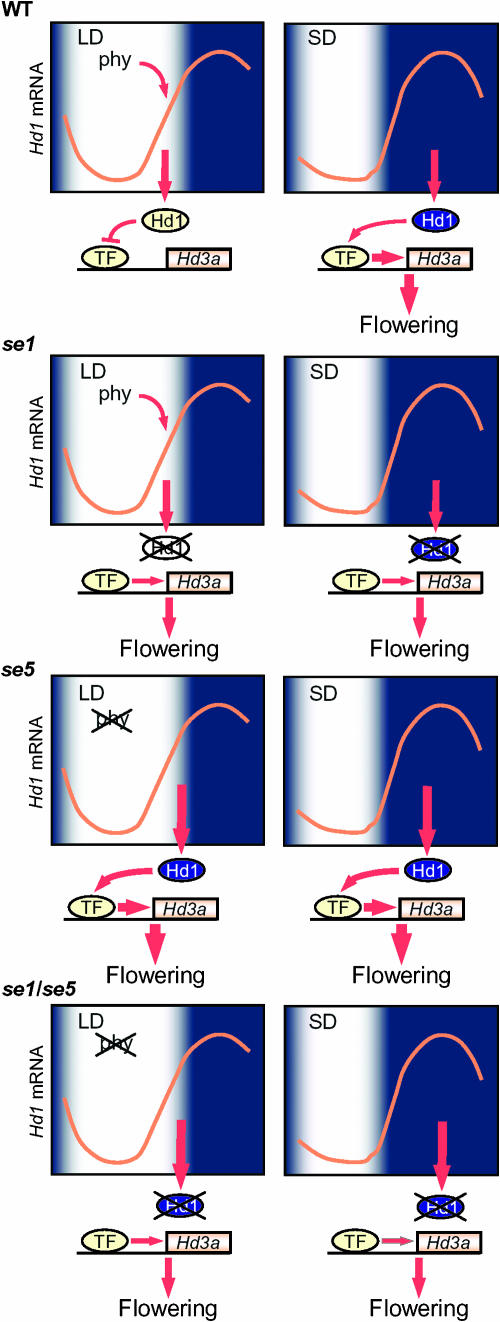
A model of daylength measurement in rice. Hd1, a CO homolog in rice, has the independent functions of inhibiting and promoting flowering under LDs and SDs, respectively. Hd1 mRNA exhibits diurnal rhythms under both SD and LD with their phases similar to those of CO in Arabidopsis. Hd1 mRNA highly accumulates at the mid to the end of the day under LD, and the coincidence of Hd1 expression and exposure to light suppresses the transcription of Hd3a and inhibits flowering under these conditions. Hd1 is proposed to inhibit Hd3a by suppressing the function of a transcription factor that autonomously activates Hd3a. Phytochrome modifies Hd1 function so that it can act to inhibit Hd3a. Without phytochrome activity, Hd1 induces Hd3a. Therefore, under SDs, when Hd1 accumulates at high levels during the night and phytochrome is inactive, Hd1 induces Hd3a and promotes flowering. The se1 mutant is deficient in Hd1 and exhibits early flowering under LDs due to the lack of Hd1 during the day. This mutant also shows late flowering under SDs due to the absence of Hd1 during the night. The se5 mutant is defective in phytochrome activity and shows early flowering irrespective of the daylength, because Hd1 is constitutively in the dark form. The double mutant shows later flowering than the se5 mutant due to the lack of the dark form of Hd1.
References
-
- Alabadi D, Oyama T, Yanovsky MJ, Harmon FG, Mas P, Kay SA (2001) Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293: 880–883 - PubMed
-
- Bünning E (1936) Die endogene Tagesrhythmik als Grundlage der photoperiodischen Reaktion. Ber Dtsch Bot Ges 54: 590–607
-
- Doyle MR, Davis SJ, Bastow RM, McWatters HG, Kozma-Bognar L, Nagy F, Millar AJ, Amasino RM (2002) The ELF4 gene controls circadian rhythms and flowering time in Arabidopsis thaliana. Nature 419: 74–77 - PubMed
-
- Dunlap JC (1999) Molecular bases for circadian clocks. Cell 96: 271–290 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
