

The Euplotes telomerase subunit p43 stimulates enzymatic activity and processivity in vitro
- PMID: 15208446
- PMCID: PMC1370601
- DOI: 10.1261/rna.7400704
The Euplotes telomerase subunit p43 stimulates enzymatic activity and processivity in vitro
Abstract
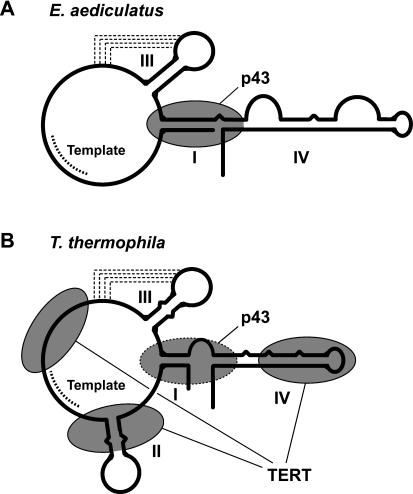
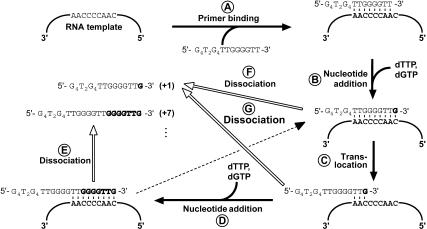
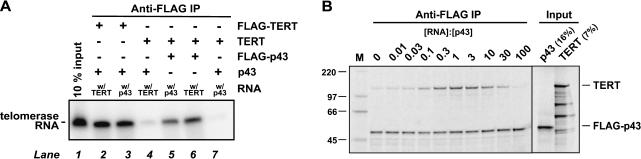
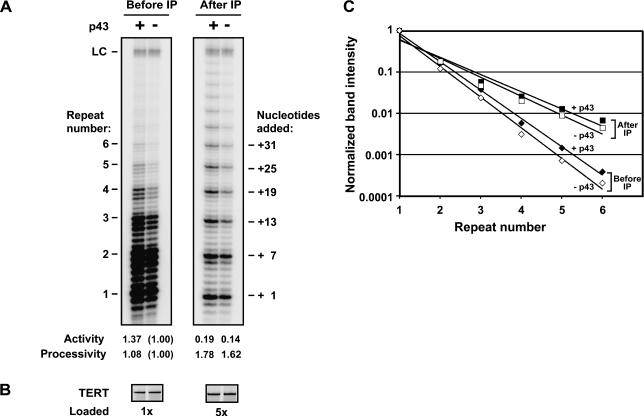
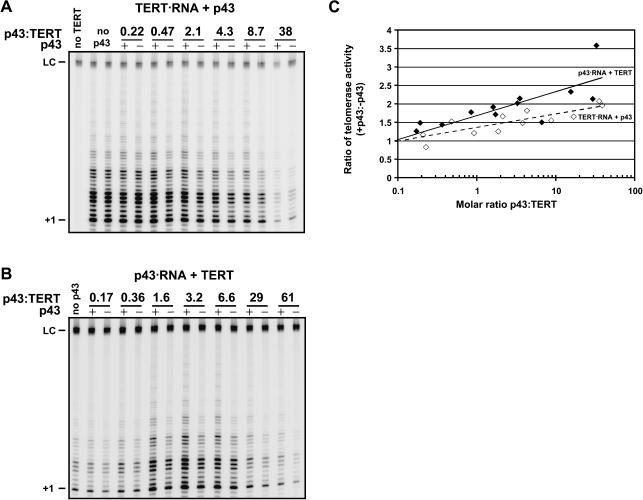
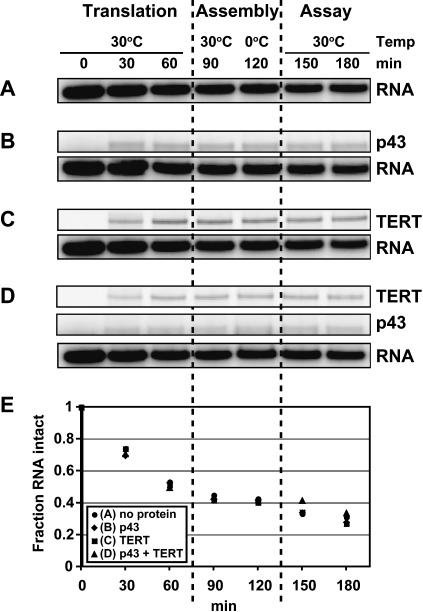
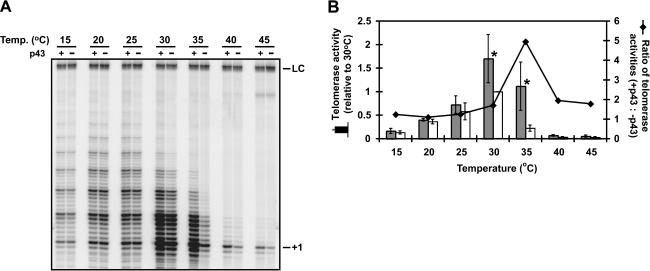
Telomerase is a reverse transcriptase that synthesizes telomeric DNA repeats at the ends of eukaryotic chromosomes. Although it is minimally composed of a conserved catalytic protein subunit (TERT) and an RNA component, additional accessory factors present in the holoenzyme play crucial roles in the biogenesis and function of the enzyme complex. Telomerase from the ciliate Tetrahymena can be reconstituted in active form in vitro. Using this system, we show that p43, a telomerase-specific La-motif protein from the ciliate Euplotes, stimulates activity and increases repeat addition processivity of telomerase. Activity enhancement by p43 requires its incorporation into a TERT.RNA.p43 ternary complex but is independent of other dissociable protein factors functioning in telomerase complex assembly. Stimulation is enhanced at elevated temperatures, supporting a role for p43 in structural stabilization of a critical region of the RNA subunit. To our knowledge, this represents the first demonstration that an authentic telomerase accessory protein can directly affect the enzymatic activity of the core enzyme in vitro.
Figures
References
-
- Aigner, S., Postberg, J., Lipps, H.J., and Cech, T.R. 2003. The Euplotes La motif protein p43 has properties of a telomerase-specific subunit. Biochemistry 42: 5736–5747. - PubMed
-
- Bryan, T.M., Goodrich, K.J., and Cech, T.R. 2000a. A mutant of Tetrahymena telomerase reverse transcriptase with increased processivity. J. Biol. Chem. 275: 24199–24207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources