

doi: 10.1042/BJ20040762.
Heparan sulphate identified on human erythrocytes: a Plasmodium falciparum receptor
Affiliations
- PMID: 15209561
- PMCID: PMC1133867
- DOI: 10.1042/BJ20040762
Item in Clipboard
Heparan sulphate identified on human erythrocytes: a Plasmodium falciparum receptor
Biochem J.
.
Abstract
HS (heparan sulphate) has hitherto not been found on human red blood cells (RBCs, erythrocytes). However, malarial-parasite (Plasmodium falciparum)-infected RBCs adhere to uninfected RBCs via HS-like receptors. In the present paper we demonstrate that human RBCs carry epitopes for an anti-HS antibody. Glycans isolated from RBC membranes reacted to HS-specific degradations and adhered to an HS-binding malaria antigen. Additionally, an HS core protein was identified. This suggests that HS is present on human RBCs.
Figures
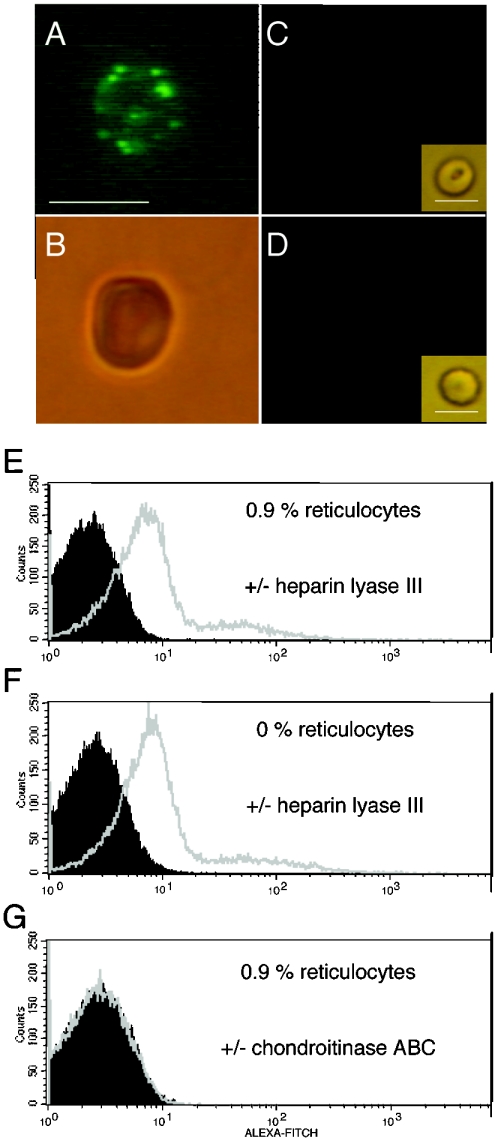
RBCs were treated with neuraminidase, and heparin lyase III or chondroitinase ABC, followed by staining with 3G10 antibody, anti-mouse FITC-labelled antibody and Alexa-Fluor-labelled anti-FITC antibody as described in the Materials and methods section. Cells were analysed by microscopy in visible light or incident UV light (A–D) and by FACS (E and G). RBCs treated with (A) neuraminidase and heparin lyase III under UV light, (B) under visible light, (C) RBCs treated with neuraminidase and chondroitinase ABC under UV light and under visible light (insert) and (D) untreated RBCs under UV light and under visible light (insert) (bar represents 6 μm) are shown. Also shown are FACS analyses (E) of RBC populations containing 0.9% reticulocytes after heparin lyase III treatment (grey) or in the absence of heparin lyase III treatment (black), (F) of a reticulocyte-free population after heparin lyase III treatment (grey) or in the absence of heparin lyase III treatment (black), and (G) of RBCs containing 0.9% reticulocytes after chondroitinase ABC treatment (grey) or in the absence of chondroitinase ABC treatment (black).
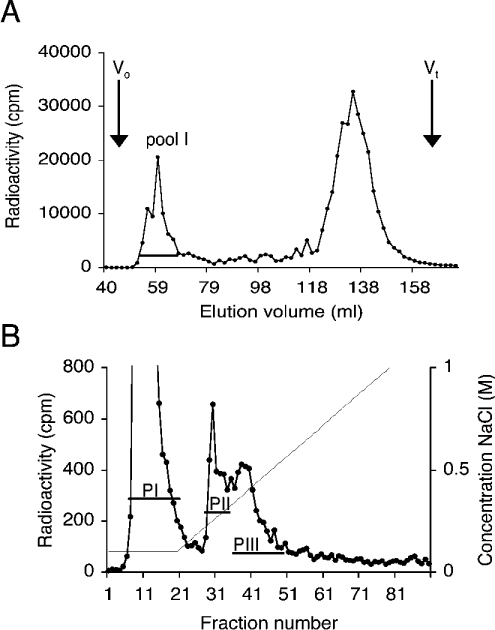
(A) Size separation of O-glycans. 3H-labelled O-glycans isolated from RBCs were separated on a Sepharose G-50 column, and fractions containing large O-glycans in pool I were collected. The void volume (Vo) and the total included volume (Vt) are indicated. (B) Anion-exchange chromatography of large O-glycans. Pool I glycans (A) were separated on a DEAE-Sephacel column and eluted with an NaCl gradient. Fractions were collected, and two peaks (PII and PIII) were pooled and analysed for HS content. Both chromatograms represent typical results of at least three different preparations.

Size separation of O-glycans (Figure 2B, PIII) on a Superose 12 column before (A) and after (B) heparin lyase III treatment, or after deamination at pH 1.5 (C). The void volume (Vo), the total included volume (Vt) and the elution positions of standard heparin oligosaccharides of the indicated sizes (26-, 16-, 10- and 2-mers) are marked. The analysis was repeated twice.
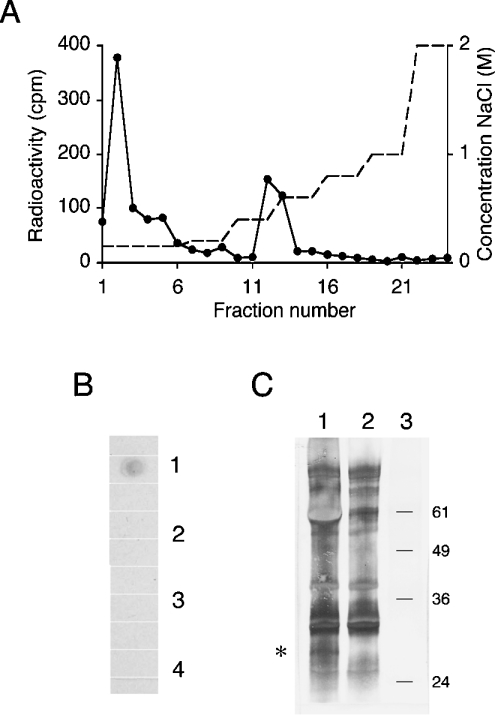
(A) O-glycans from RBCs (Figure 2B, PIII) were tested for binding to the DBL1α affinity column. Bound material was eluted with an NaCl gradient. (B) Dot-blot of RBC membrane extracts treated with (1) active heparin lyase III, (2) inactive heparin lyase III, (3) chondroitinase ABC or (4) buffer. Extracts were dotted on to a nitrocellulose membrane. The membrane was developed with 3G10 antibody and horseradish-peroxidase-conjugated anti-mouse antibody. (C) RBC membranes were isolated from biotinylated intact RBCs and treated with active heparin lyase III (lane 1) or inactive heparin lyase III (lane 2), separated by SDS/12%-(w/v)-PAGE and analysed by immunoblotting. Biotinylated proteins were detected with alkaline phosphatase-conjugated ExtrAvidin. An additional band of ≈30 kDa was detected after treatment with active heparin lyase III (asterisk). The mobility of molecular-mass standard proteins (kDa) is indicated to the right (lane 3).
References
-
- Drzeniek Z., Stöcker G., Siebertz B., Just U., Schroeder T., Ostertag W., Haubeck H.-D. Heparan sulfate proteoglycan expression is induced during early erythroid differentiation of multipotent hematopoietic stem cells. Blood. 1999;93:2884–2897. - PubMed
-
- Miller L. H., Good M. F., Milon G. Malaria pathogenesis. Science. 1994;264:1878–1883. - PubMed
-
- Baruch D. I., Pasloske B. L., Singh H. B., Bi X., Ma X. C., Feldman M., Taraschi T. F., Howard R. J. Cloning the P. falciparum gene endcoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell. 1995;82:77–87. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources