

Efficient downregulation of immunoglobulin mu mRNA with premature translation-termination codons requires the 5'-half of the VDJ exon
- PMID: 15210863
- PMCID: PMC443527
- DOI: 10.1093/nar/gkh651
Efficient downregulation of immunoglobulin mu mRNA with premature translation-termination codons requires the 5'-half of the VDJ exon
Abstract
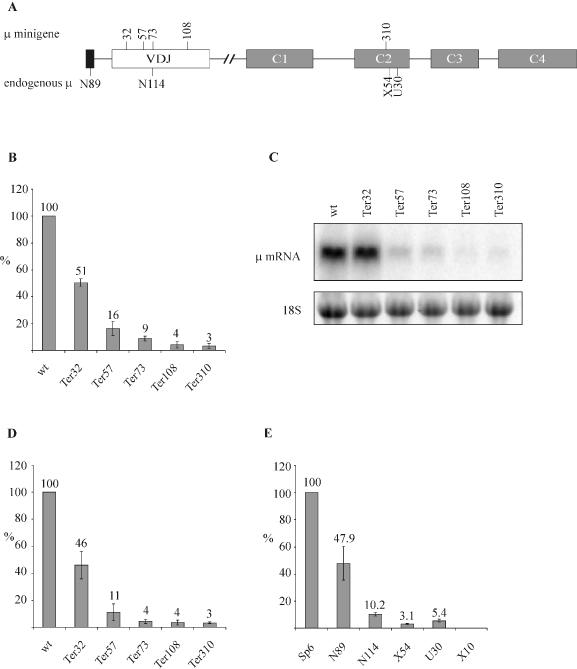
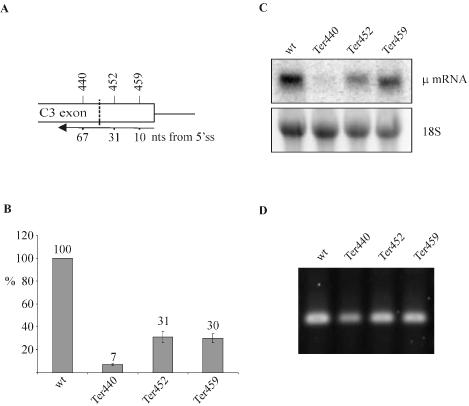
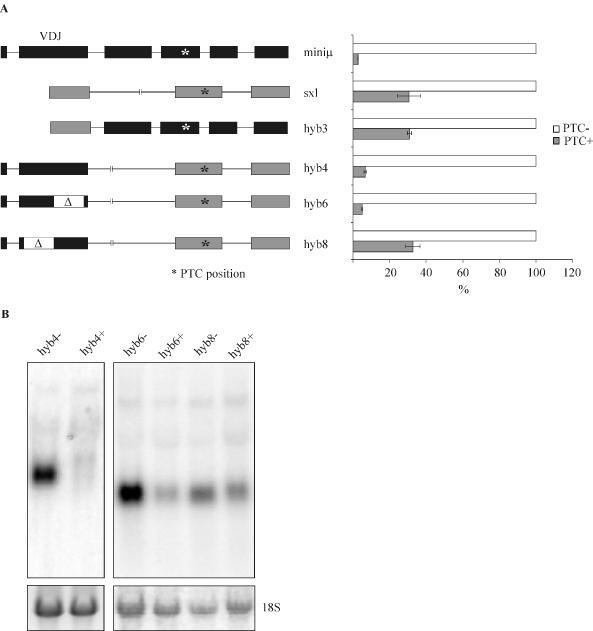
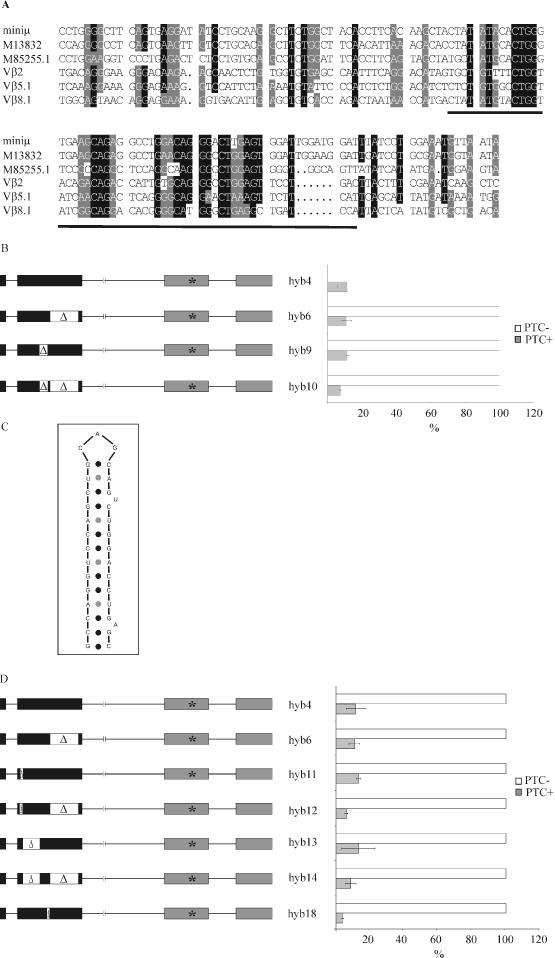
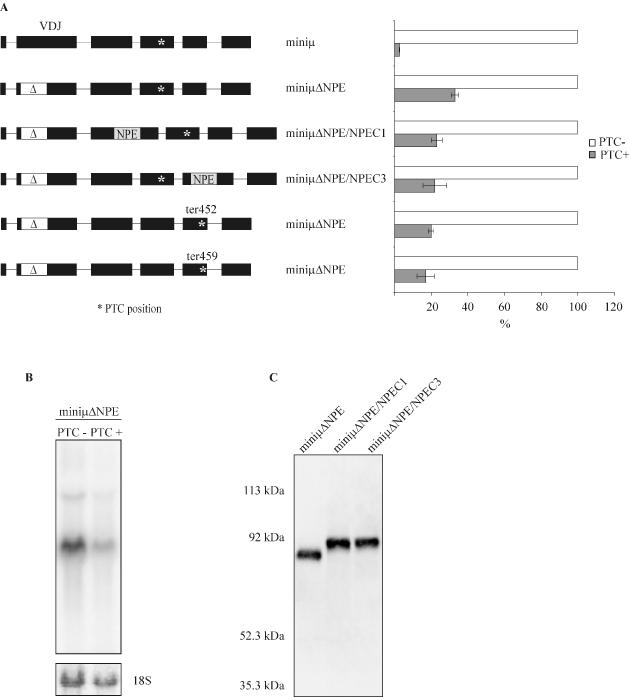
Premature translation-termination codons (PTCs) elicit rapid degradation of the mRNA by a process called nonsense-mediated mRNA decay (NMD). NMD appears to be significantly more efficient for mRNAs of genes belonging to the immunoglobulin superfamily, which frequently acquire PTCs during VDJ rearrangment, than for mRNAs of other genes. To identify determinants for efficient NMD, we developed a minigene system derived from a mouse immunoglobulin micro gene (Ig-micro) and measured the effect of PTCs at different positions on the mRNA level. This revealed that PTCs located downstream of the V-D junction in the VDJ exon of Ig-micro minigenes and of endogenous Ig-micro genes elicit very strong mRNA downregulation, whereas NMD efficiency decreases gradually further upstream in the V segment where a PTC was inserted. Interestingly, two PTCs are in positions where they usually do not trigger NMD (<50 nt from the 3'-most 5' splice site) still resulted in reduced mRNA levels. Using a set of hybrid constructs comprised of Ig-micro and an inefficient substrate for NMD, we identified a 177 nt long element in the V segment that is necessary for efficient downregulation of PTC-containing hybrid transcripts. Moreover, deletion of this NMD-promoting element from the Ig-micro minigene results in loss of strong NMD.
Figures
References
-
- Hentze M.W. and Kulozik,A.E. (1999) A perfect message: RNA surveillance and nonsense-mediated decay. Cell, 96, 307–310. - PubMed
-
- Li S. and Wilkinson,M.F. (1998) Nonsense surveillance in lymphocytes? Immunity, 8, 135–141. - PubMed
-
- Maquat L.E. (2004) Nonsense-mediated mRNA decay: splicing, translation and mRNP dynamics. Nature Rev. Mol. Cell Biol., 5, 89–99. - PubMed
-
- Mendell J.T. and Dietz,H.C. (2001) When the message goes awry. Disease-producing mutations that influence mRNA content and performance. Cell, 107, 411–414. - PubMed
-
- Wilusz C.J., Wang,W. and Peltz,S.W. (2001) Curbing the nonsense: the activation and regulation of mRNA surveillance. Genes Dev., 15, 2781–2785. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
