

The two murein lipoproteins of Salmonella enterica serovar Typhimurium contribute to the virulence of the organism
- PMID: 15213144
- PMCID: PMC427434
- DOI: 10.1128/IAI.72.7.3987-4003.2004
The two murein lipoproteins of Salmonella enterica serovar Typhimurium contribute to the virulence of the organism
Abstract
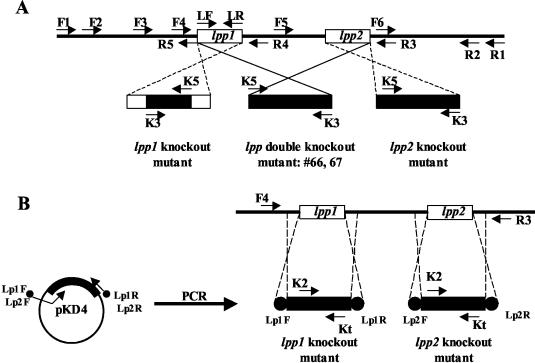
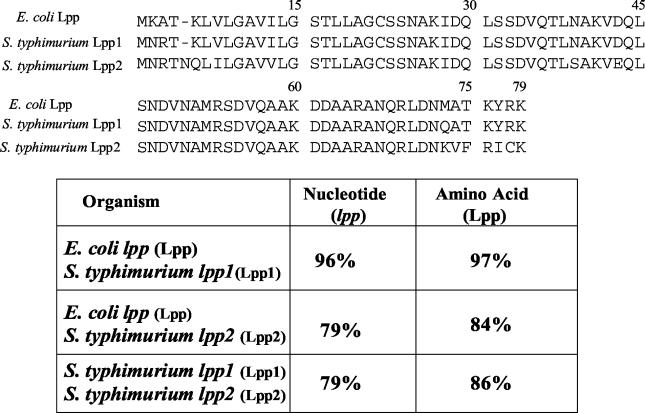
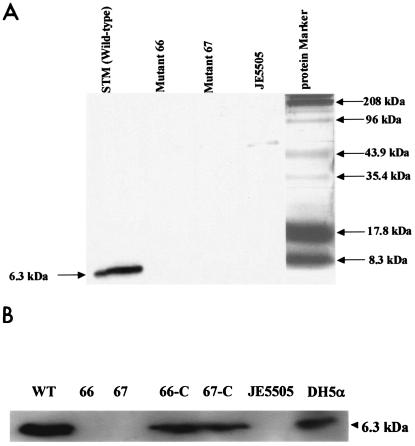
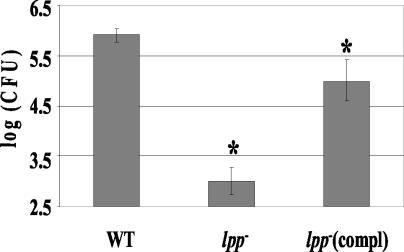
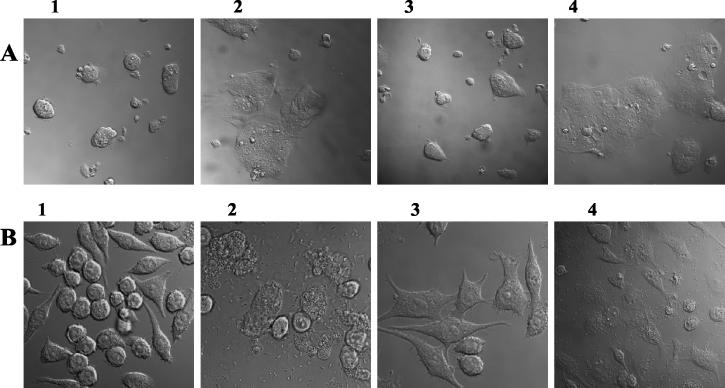
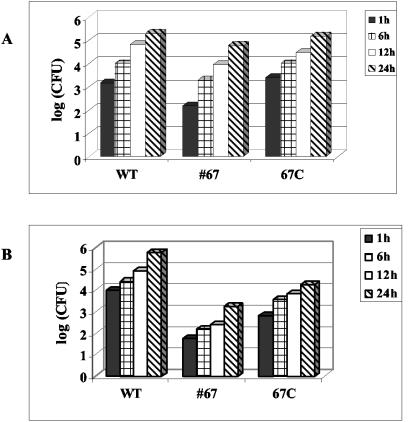
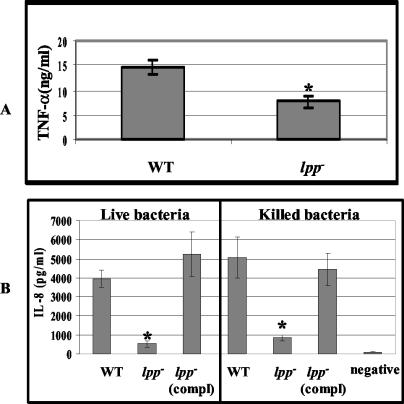
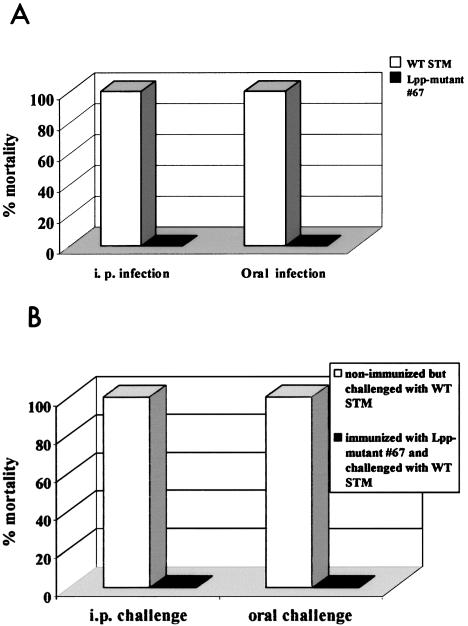
Septic shock due to Salmonella and other gram-negative enteric pathogens is a leading cause of death worldwide. The role of lipopolysaccharide in sepsis is well studied; however, the contribution of other bacterial outer membrane components, such as Braun (murein) lipoprotein (Lpp), is not well defined. The genome of Salmonella enterica serovar Typhimurium harbors two copies of the lipoprotein (lpp) gene. We constructed a serovar Typhimurium strain with deletions in both copies of the lpp gene (lpp1 and lpp2) by marker exchange mutagenesis. The integrity of the cell membrane and the secretion of the effector proteins through the type III secretion system were not affected in the lpp double-knockout mutant. Subsequently, the virulence potential of this mutant was examined in a cell culture system using T84 intestinal epithelial and RAW264.7 macrophage cell lines and a mouse model of salmonellosis. The lpp double-knockout mutant was defective in invading and inducing cytotoxic effects in T84 and RAW264.7 cells, although binding of the mutant to the host cell was not affected when compared to the wild-type (WT) serovar Typhimurium. The motility of the mutant was impaired, despite the finding that the number of flagella was similar in the lpp double knockout mutant and the WT serovar Typhimurium. Deletion in the lpp genes did not affect the intracellular survival and replication of Salmonella in macrophages and T84 cells. Induction of the proinflammatory cytokines tumor necrosis factor alpha and interleukin-8 (IL-8) was significantly reduced in macrophages and T84 cells infected with the lpp double-knockout mutant. The levels of IL-8 remained unaffected in T84 cells when infected with either live or heat-killed WT and lpp mutant, indicating that invasion was not required for IL-8 production and that Toll-like receptor 2 signaling might be affected in the Lpp double-knockout mutant. These effects of the Lpp protein could be restored by complementation of the isogenic mutant. The lpp double-knockout mutant was avirulent in mice, and animals infected with this mutant were protected from a lethal challenge dose of WT serovar Typhimurium. The severe combined immunodeficient mice, on the other hand, were susceptible to infection by the lpp double-knockout mutant. The serovar Typhimurium mutants from which only one of the lpp (lpp1 or lpp2) genes was deleted were also avirulent in mice. Taken together, our data indicated that Lpp specifically contributed to the virulence of the organism.
Figures
Similar articles
-
Murein lipoprotein is a critical outer membrane component involved in Salmonella enterica serovar typhimurium systemic infection.Infect Immun. 2005 Feb;73(2):1081-96. doi: 10.1128/IAI.73.2.1081-1096.2005. Infect Immun. 2005. PMID: 15664952 Free PMC article.
-
Global gene expression of a murein (Braun) lipoprotein mutant of Salmonella enterica serovar Typhimurium by microarray analysis.Gene. 2006 Jun 7;374:121-7. doi: 10.1016/j.gene.2006.01.034. Epub 2006 Mar 29. Gene. 2006. PMID: 16574345
-
Protective Immunity Elicited by Oral Immunization of Mice with Salmonella enterica Serovar Typhimurium Braun Lipoprotein (Lpp) and Acetyltransferase (MsbB) Mutants.Front Cell Infect Microbiol. 2016 Nov 10;6:148. doi: 10.3389/fcimb.2016.00148. eCollection 2016. Front Cell Infect Microbiol. 2016. PMID: 27891321 Free PMC article.
-
Salmonella enterica serovar Typhimurium skills to succeed in the host: virulence and regulation.Clin Microbiol Rev. 2013 Apr;26(2):308-41. doi: 10.1128/CMR.00066-12. Clin Microbiol Rev. 2013. PMID: 23554419 Free PMC article. Review.
-
Regulation and function of the Salmonella MgtC virulence protein.J Microbiol. 2015 Oct;53(10):667-72. doi: 10.1007/s12275-015-5283-1. Epub 2015 Aug 1. J Microbiol. 2015. PMID: 26231375 Review.
Cited by
-
A Biological Signature for the Inhibition of Outer Membrane Lipoprotein Biogenesis.mBio. 2022 Jun 28;13(3):e0075722. doi: 10.1128/mbio.00757-22. Epub 2022 Jun 13. mBio. 2022. PMID: 35695460 Free PMC article.
-
Deletion of the Braun lipoprotein-encoding gene and altering the function of lipopolysaccharide attenuate the plague bacterium.Infect Immun. 2013 Mar;81(3):815-28. doi: 10.1128/IAI.01067-12. Epub 2012 Dec 28. Infect Immun. 2013. PMID: 23275092 Free PMC article.
-
During infection of epithelial cells Salmonella enterica serovar Typhimurium undergoes a time-dependent transcriptional adaptation that results in simultaneous expression of three type 3 secretion systems.Cell Microbiol. 2008 Apr;10(4):958-84. doi: 10.1111/j.1462-5822.2007.01099.x. Epub 2007 Nov 20. Cell Microbiol. 2008. PMID: 18031307 Free PMC article.
-
Attenuation of Salmonella enterica Serovar Typhimurium by altering biological functions of murein lipoprotein and lipopolysaccharide.Infect Immun. 2005 Dec;73(12):8433-6. doi: 10.1128/IAI.73.12.8433-8436.2005. Infect Immun. 2005. PMID: 16299345 Free PMC article.
-
Cold shock exoribonuclease R (VacB) is involved in Aeromonas hydrophila pathogenesis.J Bacteriol. 2008 May;190(10):3467-74. doi: 10.1128/JB.00075-08. Epub 2008 Mar 14. J Bacteriol. 2008. PMID: 18344363 Free PMC article.
References
-
- Aliprantis, A. O., R. B. Yang, M. R. Mark, S. Suggett, B. Devaux, J. D. Radolf, G. R. Klimpel, P. Godowski, and A. Zychlinsky. 1999. Cell activation and apoptosis by bacterial lipoproteins through toll-like receptor-2. Science 285:736-739. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. Seidman, J. A. Smith, and K. Struhl. 1989. Current protocols in molecular biology. John Wiley and Sons, Inc., New York, N.Y.
-
- Babu, M. M., and K. Sankaran. 2002. DOLOP—database of bacterial lipoproteins. Bioinformatics 18:641-643. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
