

Nuclear factor-kappaB regulates inflammatory cell apoptosis and phagocytosis in rat carrageenin-sponge implant model
- PMID: 15215167
- PMCID: PMC1618533
- DOI: 10.1016/s0002-9440(10)63280-4
Nuclear factor-kappaB regulates inflammatory cell apoptosis and phagocytosis in rat carrageenin-sponge implant model
Abstract
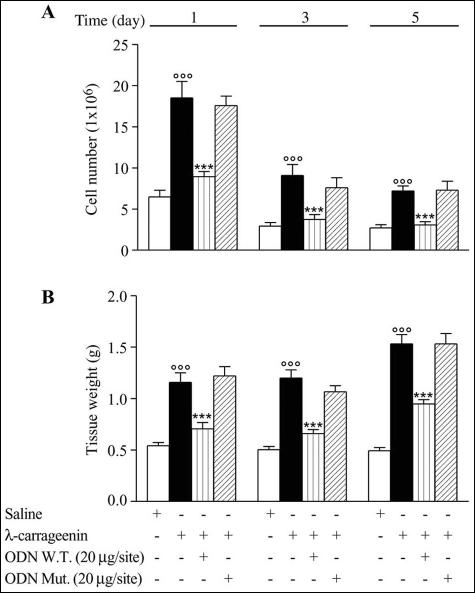
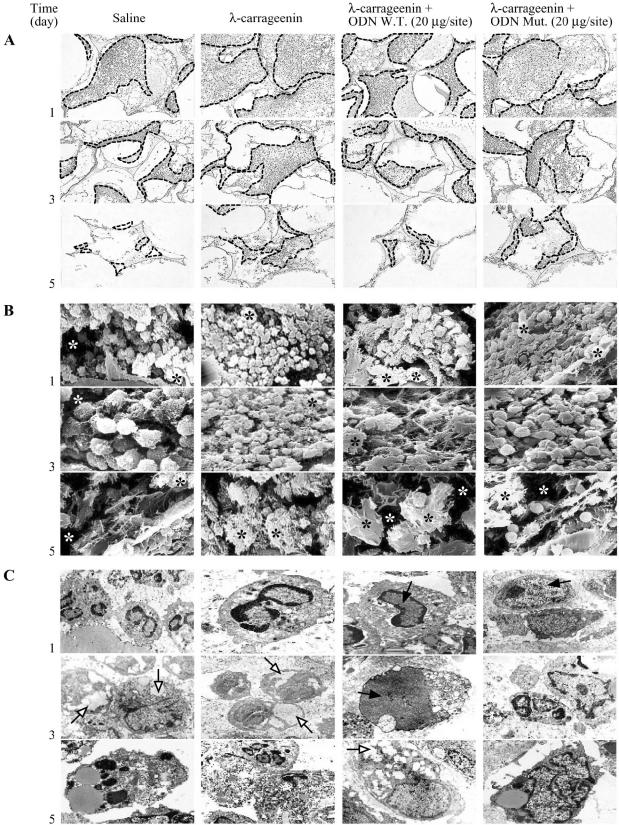
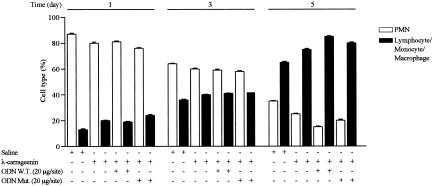
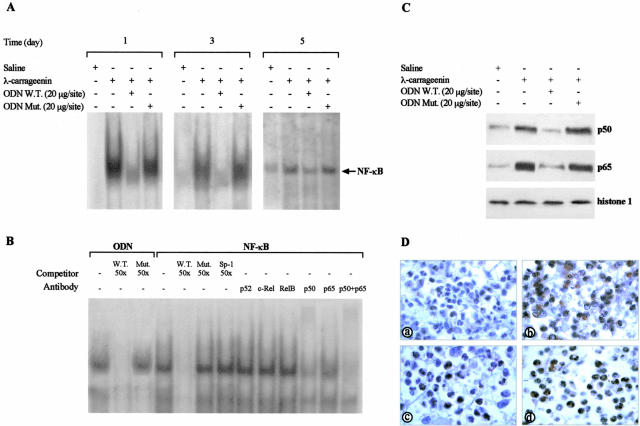
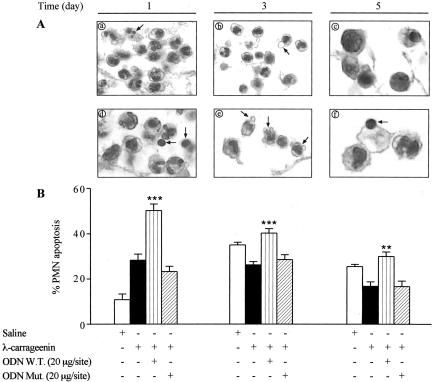
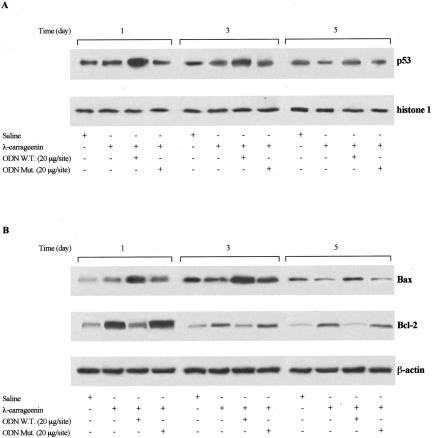
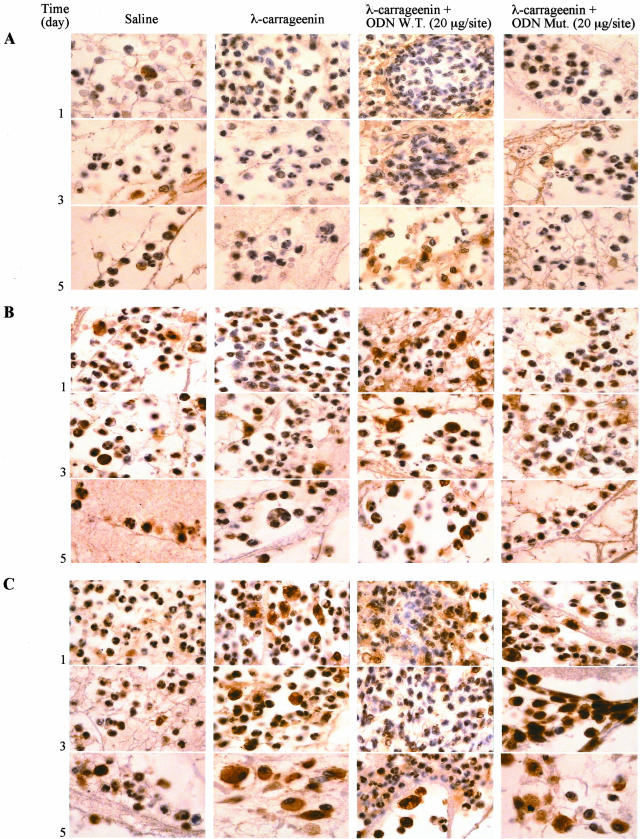
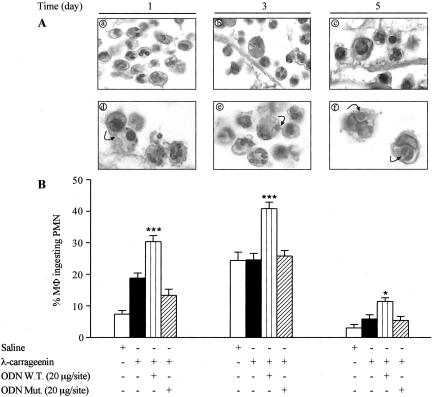
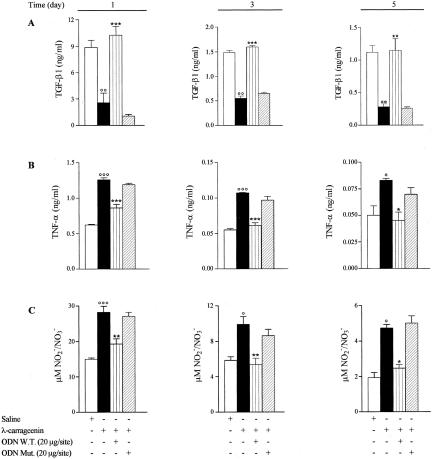
In the present study we investigated whether apoptosis and phagocytosis are regulated by nuclear factor (NF)-kappaB in a model of chronic inflammation. The subcutaneous implant of lambda-carrageenin-soaked sponges elicited an inflammatory response, characterized by a time-related increase of leukocyte infiltration into the sponge and tissue formation, which was inhibited by simultaneous injection of wild-type oligodeoxynucleotide decoy to NF-kappaB. Molecular and morphological analysis performed on infiltrated cells demonstrated: 1) an inhibition of NF-kappaB/DNA binding activity; 2) an increase of polymorphonuclear leukocyte apoptosis correlated either to an increase of p53 or Bax and decrease of Bcl-2 protein expression; and 3) an increase of phagocytosis of apoptotic polymorphonuclear leukocytes by macrophages associated with an increase of transforming growth factor-beta1 and decrease of tumor necrosis factor-alpha as well as nitrite/nitrate production. Our results, showing that blockade of NF-kappaB by oligodeoxynucleotide decoy increases inflammatory cell apoptosis and phagocytosis, may contribute to lead to new insights into the mechanisms governing the inflammatory process.
Figures
References
-
- Savill J. Apoptosis in resolution of inflammation. J Leukoc Biol. 1997;61:375–380. - PubMed
-
- Kobayashi SD, Voyich JM, Somerville JA, Braughton KR, Malech HL, Musser JM, DeLeo FR. An apoptosis-differentiation program in human polymorphonuclear leukocytes facilitates resolution of inflammation. J Leukoc Biol. 2003;73:315–322. - PubMed
-
- Savill J, Dransfield I, Gregory C, Haslett C. A blast from the past: clearance of apoptotic cells regulates immune responses. Nat Rev. 2002;2:965–975. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
