

Regulation of macrophage foam cell formation by alphaVbeta3 integrin: potential role in human atherosclerosis
- PMID: 15215180
- PMCID: PMC1618536
- DOI: 10.1016/s0002-9440(10)63293-2
Regulation of macrophage foam cell formation by alphaVbeta3 integrin: potential role in human atherosclerosis
Abstract
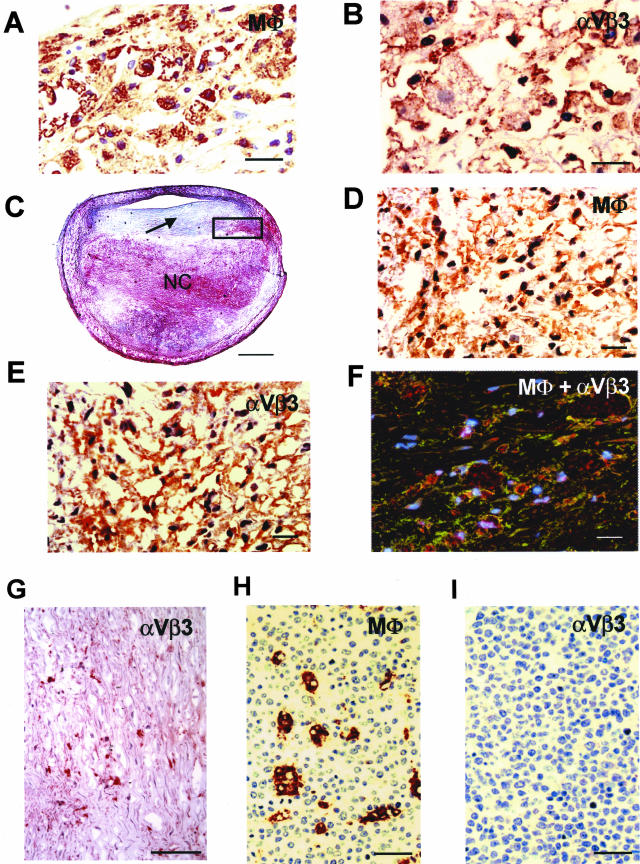
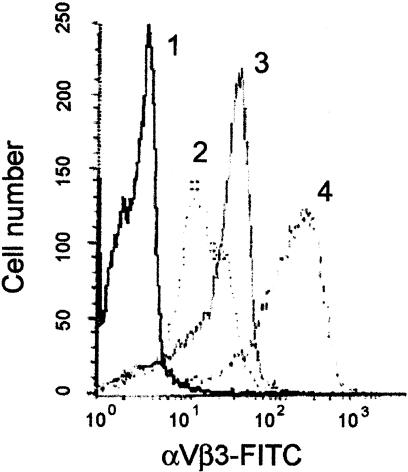
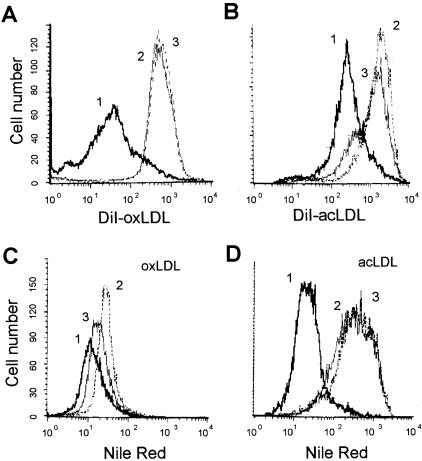
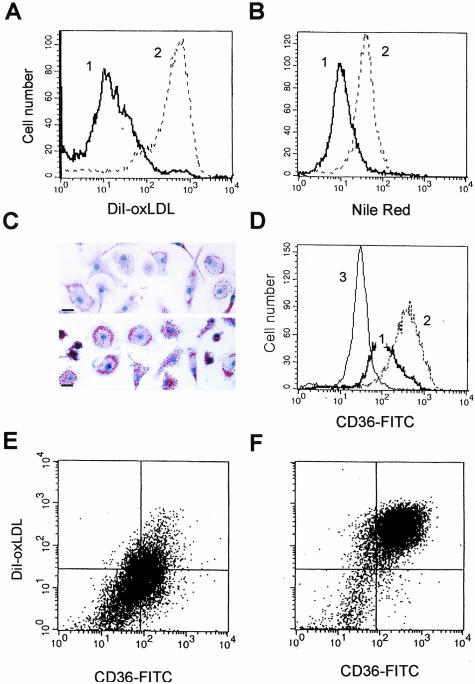
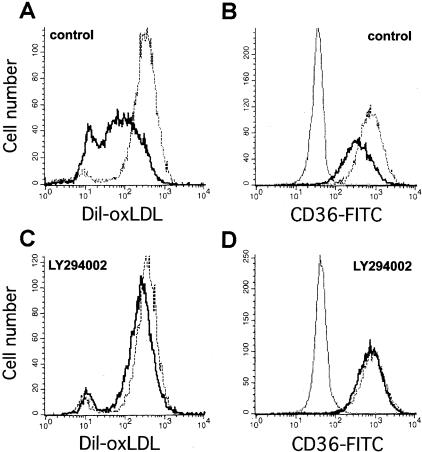
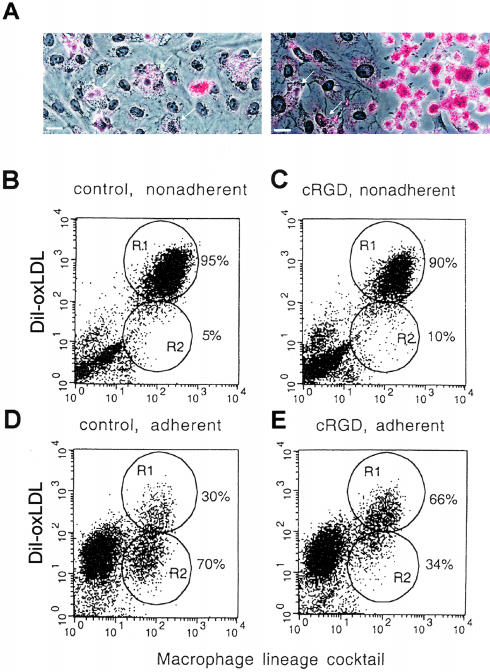
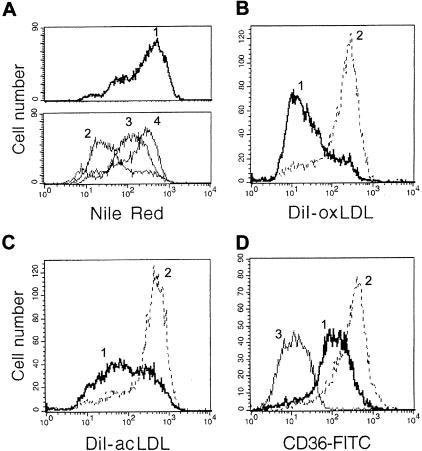
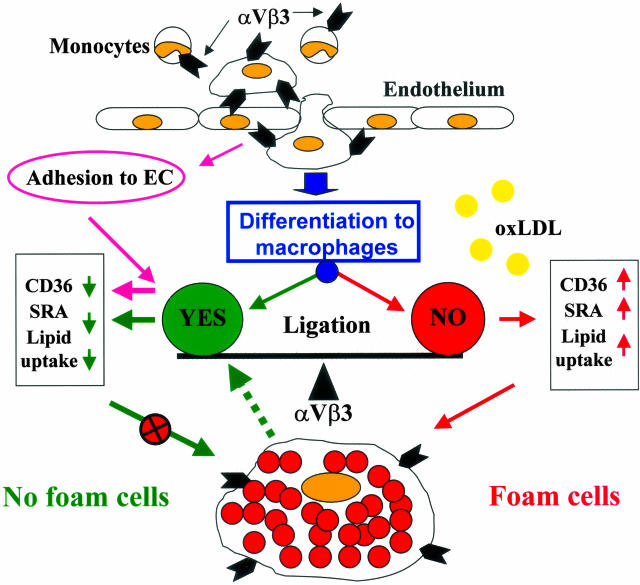
The accumulation of macrophage foam cells in atherosclerotic lesions is associated with both initiation and progression of this disease. Scavenger receptors CD36 and SRA are the primary receptors responsible for conversion of macrophages into foam cells. Integrin alphaVbeta3 plays a role in the differentiation of several cell types, but its involvement in the transition of macrophages into foam cells and the potential role of this receptor in atherosclerosis have not been examined. Using an in vitro model of single surface receptor activation by binding with an immobilized monoclonal antibody specific to alphaVbeta3 integrin we show that ligation of alphaVbeta3 integrin prevents differentiation of blood monocytes and macrophages into the foam cell phenotype via coordinate down-regulation of CD36 and SRA. This effect of alphaVbeta3 integrin ligation can be reproduced by contact with endothelial cells, whereas the inhibition of alphaVbeta3 receptor ligation restores the uptake of oxidized low-density lipoprotein. Moreover, we found that alphaVbeta3 integrin is readily detected in situ on macrophages in early and advanced atherosclerotic lesions and that in vitro exposure to oxidized low-density lipoprotein up-regulates alphaVbeta3 integrin expression. We hypothesize that alphaVbeta3 integrin regulates macrophage functional maturation into foam cells in a persistent manner, and therefore, by targeting alphaVbeta3 receptor it could potentially be possible to regulate progression of atherosclerosis in humans.
Figures
References
-
- Steinberg D, Parthasarathy S, Carew TE, Khoo JC, Witztum JL. Beyond cholesterol: modification of low density lipoproteins that increase its atherogeneity. N Engl J Med. 1989;320:915–924. - PubMed
-
- Brown MS, Goldstein JL. Atherosclerosis: scavenging for receptors. Nature. 1990;343:508–509. - PubMed
-
- Ross R. The pathogenesis of atherosclerosis: a perspective for the 1990s. Nature. 1993;362:801–809. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
