

In vivo and in vitro characterization of insulin-producing cells obtained from murine bone marrow
- PMID: 15220196
- PMCID: PMC3422216
- DOI: 10.2337/diabetes.53.7.1721
In vivo and in vitro characterization of insulin-producing cells obtained from murine bone marrow
Abstract
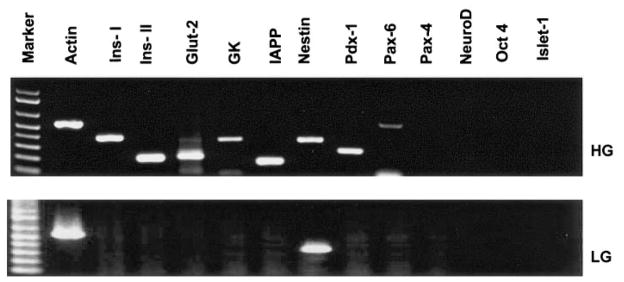
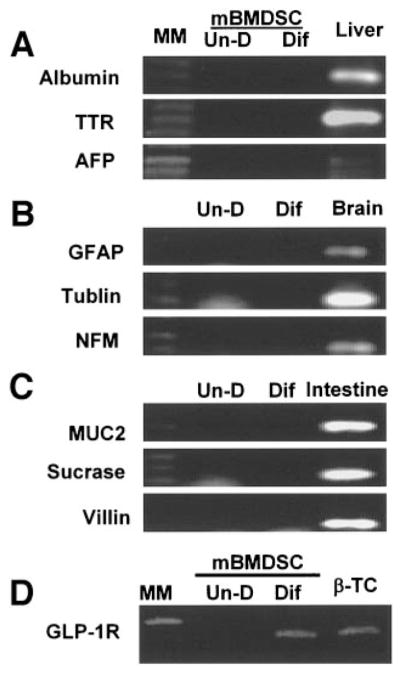
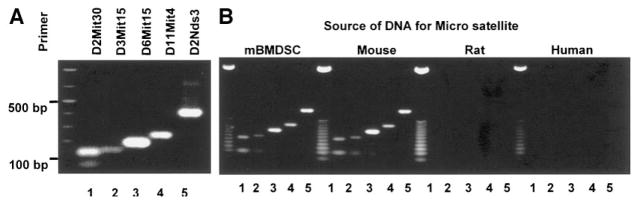
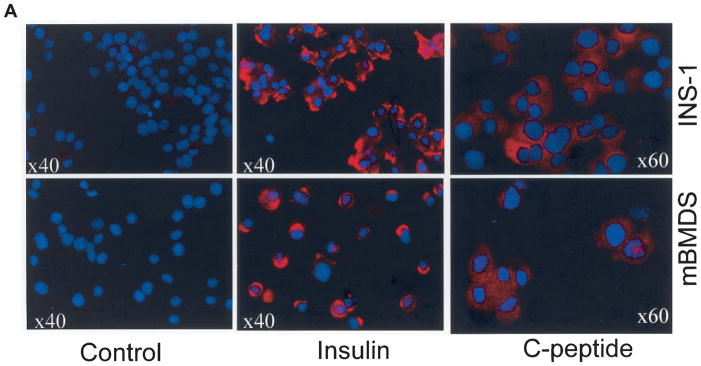
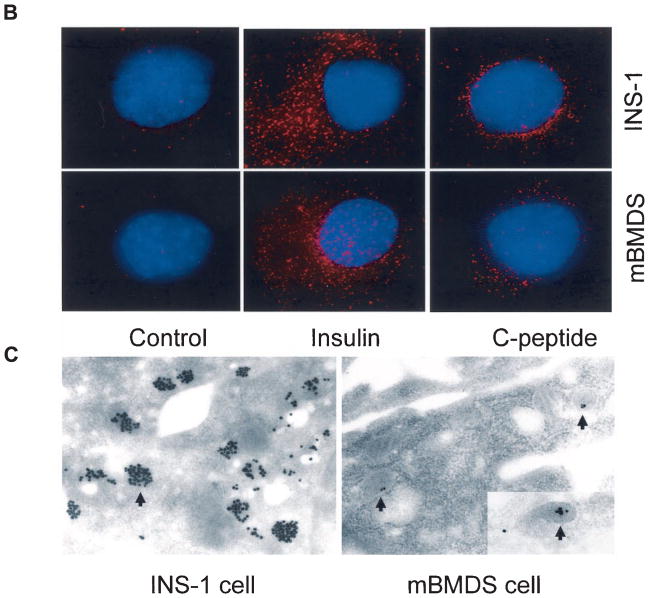
Efforts toward routine islet cell transplantation as a means for reversing type 1 diabetes have been hampered by islet availability as well as allograft rejection. In vitro transdifferentiation of mouse bone marrow (BM)-derived stem (mBMDS) cells into insulin-producing cells could provide an abundant source of autologous cells for this procedure. For this study, we isolated and characterized single cell-derived stem cell lines obtained from mouse BM. In vitro differentiation of these mBMDS cells resulted in populations meeting a number of criteria set forth to define functional insulin-producing cells. Specifically, the mBMDS cells expressed multiple genes related to pancreatic beta-cell development and function (insulin I and II, Glut2, glucose kinase, islet amyloid polypeptide, nestin, pancreatic duodenal homeobox-1 [PDX-1], and Pax6). Insulin and C-peptide production was identified by immunocytochemistry and confirmed by electron microscopy. In vitro studies involving glucose stimulation identified glucose-stimulated insulin release. Finally, these mBMDS cells transplanted into streptozotocin-induced diabetic mice imparted reversal of hyperglycemia and improved metabolic profiles in response to intraperitoneal glucose tolerance testing. These results indicate that mouse BM harbors cells capable of in vitro transdifferentiating into functional insulin-producing cells and support efforts to derive such cells in humans as a means to alleviate limitations surrounding islet cell transplantation.
Figures
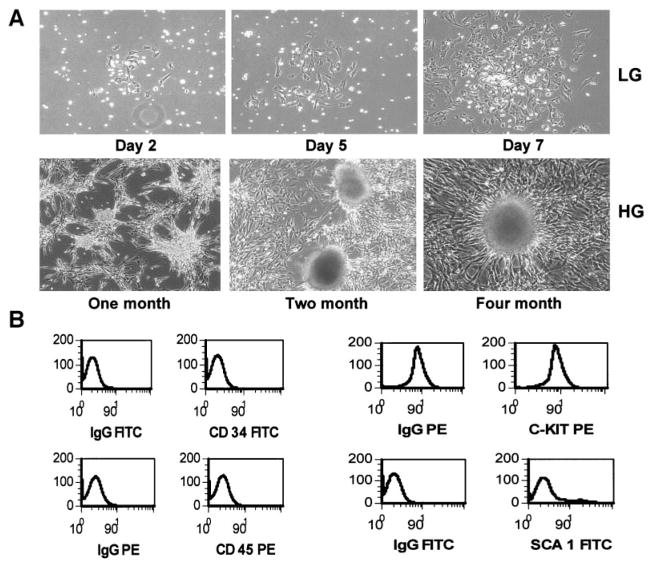
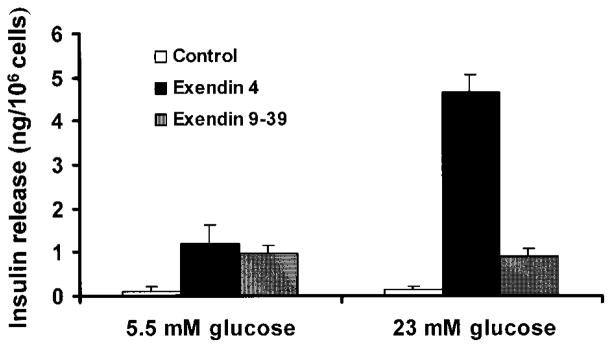
 ) upon glucose stimulation. Cells were cultured in RPMI 1640 medium containing 5.5 mmol/l glucose and 5% FCS, plus nicotinamide for 1 week and for an additional 1 week in the presence of either exendin 4 or exendin 9–39. The cells were then switched to serum-free medium containing 0.5% BSA for 12 h and then stimulated with 23 mmol/l glucose for 2 h. The cell culture medium was then collected for assay of insulin release. Released insulin in the media was detected by an ultrasensitive ELISA kit. Results shown here represent those of three separate experiments.
) upon glucose stimulation. Cells were cultured in RPMI 1640 medium containing 5.5 mmol/l glucose and 5% FCS, plus nicotinamide for 1 week and for an additional 1 week in the presence of either exendin 4 or exendin 9–39. The cells were then switched to serum-free medium containing 0.5% BSA for 12 h and then stimulated with 23 mmol/l glucose for 2 h. The cell culture medium was then collected for assay of insulin release. Released insulin in the media was detected by an ultrasensitive ELISA kit. Results shown here represent those of three separate experiments.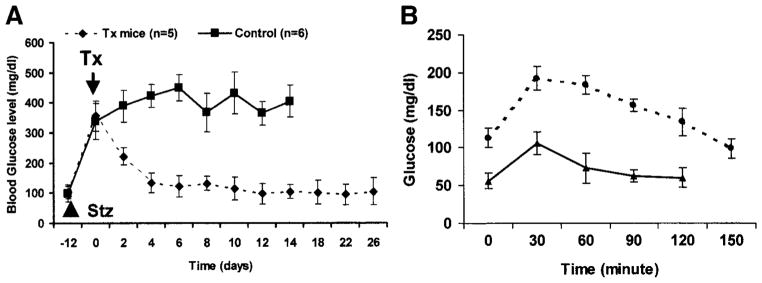
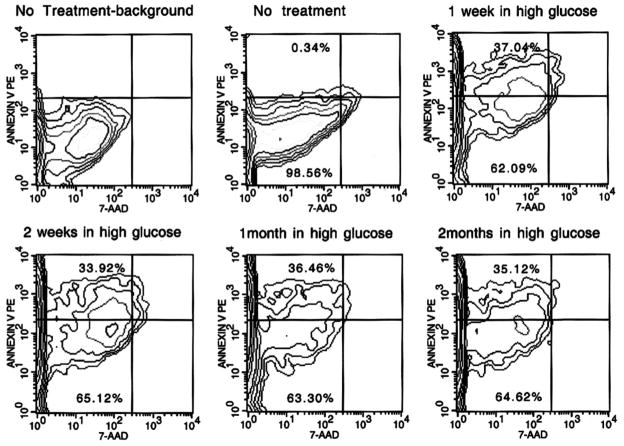
References
-
- Atkinson MA, Eisenbarth GS. Type 1 diabetes: new perspectives on disease pathogenesis and treatment. Lancet. 2001;358:221–229. - PubMed
-
- Shapiro AM, Lakey JR, Ryan EA, Korbutt GS, Toth E, Warnock GL, Kneteman NM, Rajotte RV. Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen. N Engl J Med. 2000;343:230–238. - PubMed
-
- Gunnarsson R, Klintmalm G, Lundgren G, Wilczek H, Ostman J, Groth CG. Deterioration in glucose metabolism in pancreatic transplant recipients given cyclosporin. Lancet. 1983;2:571–572. - PubMed
-
- Asahara T, Murohara T, Sullivan A, Silver M, van der ZR, Li T, Witzen-bichler B, Schatteman G, Isner JM. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275:964–967. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
