

Binding of salivary agglutinin to IgA
- PMID: 15228387
- PMCID: PMC1134054
- DOI: 10.1042/BJ20040265
Binding of salivary agglutinin to IgA
Abstract
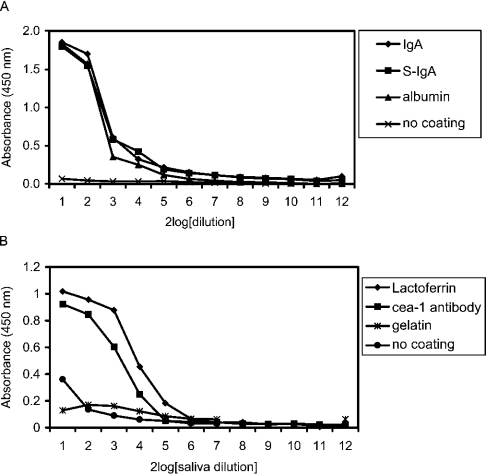
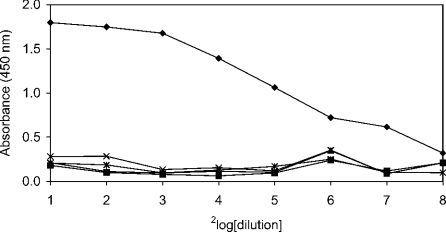
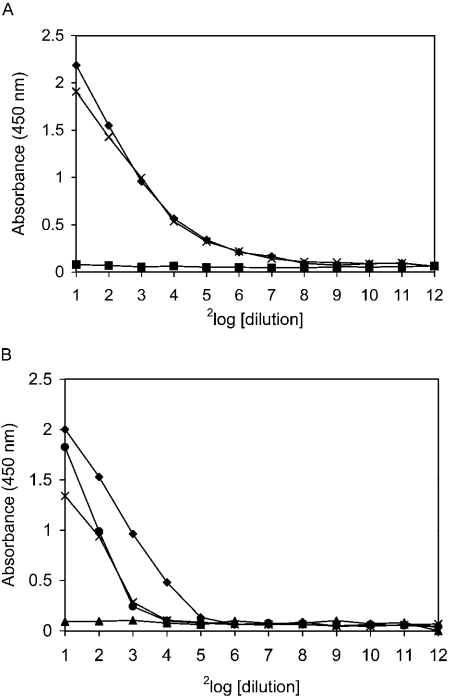
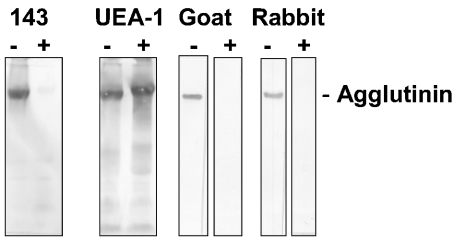
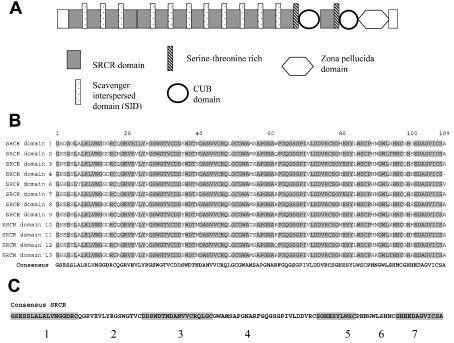
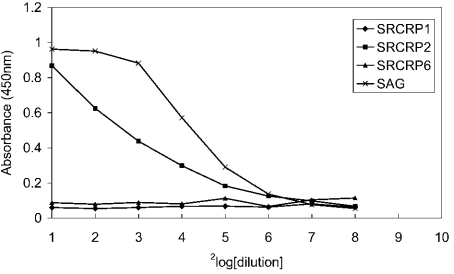
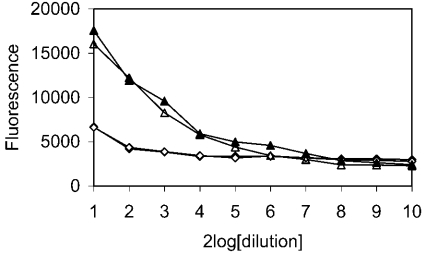
SAG (salivary agglutinin), which is identical to gp-340 (glycoprotein-340) from the lung, is encoded by DMBT1 (deleted in malignant brain tumours 1). It is a member of the SRCR (scavenger receptor cysteine-rich) superfamily and contains 14 SRCR domains, 13 of which are highly similar. SAG in saliva is partially complexed with IgA, which may be necessary for bacterial binding. The goal of the present study was to characterize the binding of purified SAG to IgA. SAG binds to a variety of proteins, including serum and secretory IgA, alkaline phosphatase-conjugated IgGs originating from rabbit, goat, swine and mouse, and lactoferrin and albumin. Binding of IgA to SAG is calcium dependent and is inhibited by 0.5 M KCl, suggesting that electrostatic interactions are involved. Binding of IgA was destroyed after reduction of SAG, suggesting that the protein moiety is involved in binding. To pinpoint further the binding domain for IgA on SAG, a number of consensus-based peptides of the SRCR domains and SRCR interspersed domains were designed and synthesized. ELISA binding studies with IgA indicated that only one of the peptides tested, comprising amino acids 18-33 (QGRVEVLYRGSWGTVC) of the 109-amino-acid SRCR domain, exhibited binding to IgA. This domain is identical to the domain of SAG that is involved in binding to bacteria. Despite this similar binding site, IgA did not inhibit binding of Streptococcus mutans to SAG or peptide. These results show that the binding of IgA to SAG is specifically mediated by a peptide sequence on the SRCR domains.
Figures
Similar articles
-
Deleted in malignant brain tumors-1 protein (DMBT1): a pattern recognition receptor with multiple binding sites.Int J Mol Sci. 2010;11(12):5212-33. doi: 10.3390/ijms1112521. Epub 2010 Dec 17. Int J Mol Sci. 2010. PMID: 21614203 Free PMC article. Review.
-
A common binding motif for various bacteria of the bacteria-binding peptide SRCRP2 of DMBT1/gp-340/salivary agglutinin.Biol Chem. 2008 Sep;389(9):1193-200. doi: 10.1515/BC.2008.135. Biol Chem. 2008. PMID: 18713006
-
Identification of the bacteria-binding peptide domain on salivary agglutinin (gp-340/DMBT1), a member of the scavenger receptor cysteine-rich superfamily.J Biol Chem. 2002 Aug 30;277(35):32109-15. doi: 10.1074/jbc.M203788200. Epub 2002 Jun 5. J Biol Chem. 2002. PMID: 12050164
-
Bacteria binding by DMBT1/SAG/gp-340 is confined to the VEVLXXXXW motif in its scavenger receptor cysteine-rich domains.J Biol Chem. 2004 Nov 12;279(46):47699-703. doi: 10.1074/jbc.M406095200. Epub 2004 Sep 7. J Biol Chem. 2004. PMID: 15355985
-
Salivary agglutinin/glycoprotein-340/DMBT1: a single molecule with variable composition and with different functions in infection, inflammation and cancer.Biol Chem. 2007 Dec;388(12):1275-89. doi: 10.1515/BC.2007.158. Biol Chem. 2007. PMID: 18020944 Review.
Cited by
-
Innate immunity glycoprotein gp-340 variants may modulate human susceptibility to dental caries.BMC Infect Dis. 2007 Jun 11;7:57. doi: 10.1186/1471-2334-7-57. BMC Infect Dis. 2007. PMID: 17562017 Free PMC article.
-
Deleted in Malignant Brain Tumors 1 (DMBT1) is present in hyaline membranes and modulates surface tension of surfactant.Respir Res. 2007 Oct 1;8(1):69. doi: 10.1186/1465-9921-8-69. Respir Res. 2007. PMID: 17908325 Free PMC article.
-
Salivary Scavenger and Agglutinin (SALSA) Is Expressed in Mucosal Epithelial Cells and Decreased in Bronchial Epithelium of Asthmatic Horses.Front Vet Sci. 2019 Nov 29;6:418. doi: 10.3389/fvets.2019.00418. eCollection 2019. Front Vet Sci. 2019. PMID: 31850379 Free PMC article.
-
Deleted in malignant brain tumors-1 protein (DMBT1): a pattern recognition receptor with multiple binding sites.Int J Mol Sci. 2010;11(12):5212-33. doi: 10.3390/ijms1112521. Epub 2010 Dec 17. Int J Mol Sci. 2010. PMID: 21614203 Free PMC article. Review.
-
Functions of cell surface-anchored antigen I/II family and Hsa polypeptides in interactions of Streptococcus gordonii with host receptors.Infect Immun. 2005 Oct;73(10):6629-38. doi: 10.1128/IAI.73.10.6629-6638.2005. Infect Immun. 2005. PMID: 16177339 Free PMC article.
References
-
- Mollenhauer J., Wiemann S., Scheurlen W., Korn B., Hayashi Y., Wilgenbus K. K., von Deimling A., Poustka A. DMBT1, a new member of the SRCR superfamily, on chromosome 10q25.3–26.1 is deleted in malignant brain tumours. Nat. Genet. 1997;17:32–39. - PubMed
-
- Prakobphol A., Xu F., Hoang V. M., Larsson T., Bergstrom J., Johansson I., Frangsmyr L., Holmskov U., Leffler H., Nilsson C., et al. Salivary agglutinin, which binds Streptococcus mutans and Helicobacter pylori, is the lung scavenger receptor cysteine-rich protein gp-340. J. Biol. Chem. 2000;275:39860–39866. - PubMed
-
- Kristiansen M., Graversen J. H., Jacobsen C., Sonne O., Hoffman H. J., Law S. K., Moestrup S. K. Identification of the haemoglobin scavenger receptor. Nature (London) 2001;409:198–201. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous