

Conserved MYC transcription factors play a key role in jasmonate signaling both in tomato and Arabidopsis
- PMID: 15231736
- PMCID: PMC443520
- DOI: 10.1101/gad.297704
Conserved MYC transcription factors play a key role in jasmonate signaling both in tomato and Arabidopsis
Abstract
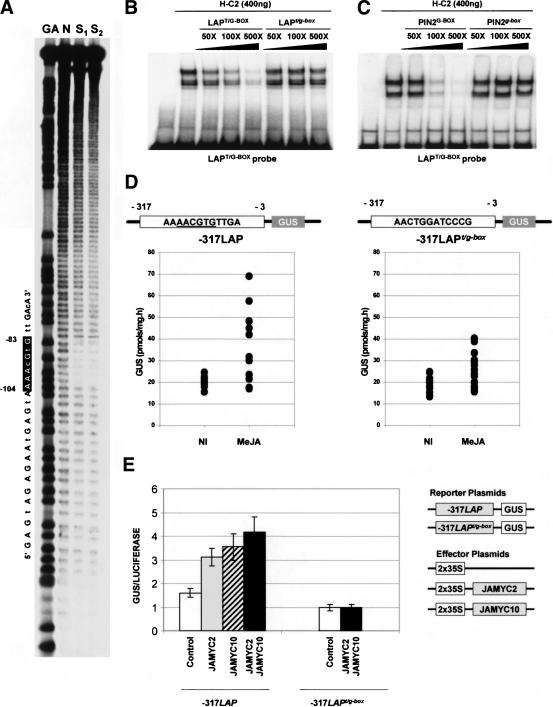
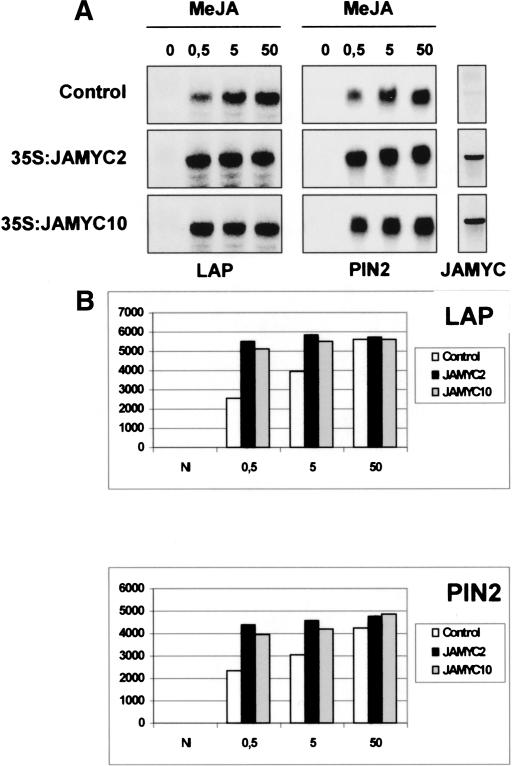
Jasmonates (JA) are important regulators of plant defense responses that activate expression of many wound-induced genes including the tomato proteinase inhibitor II (pin2) and leucine aminopeptidase (LAP) genes. Elements required for JA induction of the LAP gene are all present in the -317 to -78 proximal promoter region. Using yeast one-hybrid screening, we have identified the bHLH-leu zipper JAMYC2 and JAMYC10 proteins, specifically recognizing a T/G-box AACGTG motif in this promoter fragment. Mutation of the G-box element decreases JA-responsive LAP promoter expression. Expression of JAMYC2 and JAMYC10 is induced by JA, with a kinetics that precedes that of the LAP or pin2 transcripts. JAMYC overexpression enhanced JA-induced expression of these defense genes in potato, but did not result in constitutive transcript accumulation. Using footprinting assays, an additional protected element was identified, located directly adjacent to the T/G-box motif. Mutation of this element abolishes JA response, showing that recognition of this duplicated element is also required for gene expression. Knockout mutants in the AtMYC2 homolog gene of Arabidopsis are insensitive to JA and exhibit a decreased activation of the JA-responsive genes AtVSP and JR1. Activation of the PDF1.2 and b-CHI, ethylene/JA-responsive genes, is, however, increased in these mutants. These results show that the JAMYC/AtMYC2 transcription factors function as members of a MYC-based regulatory system conserved in dicotyledonous plants with a key role in JA-induced defense gene activation.
Figures





Similar articles
-
JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis.Plant Cell. 2004 Jul;16(7):1938-50. doi: 10.1105/tpc.022319. Epub 2004 Jun 18. Plant Cell. 2004. PMID: 15208388 Free PMC article.
-
The Arabidopsis JAZ2 promoter contains a G-Box and thymidine-rich module that are necessary and sufficient for jasmonate-dependent activation by MYC transcription factors and repression by JAZ proteins.Plant Cell Physiol. 2012 Feb;53(2):330-43. doi: 10.1093/pcp/pcr178. Epub 2011 Dec 14. Plant Cell Physiol. 2012. PMID: 22173100
-
Characterization of JAZ-interacting bHLH transcription factors that regulate jasmonate responses in Arabidopsis.J Exp Bot. 2011 Mar;62(6):2143-54. doi: 10.1093/jxb/erq408. Epub 2011 Feb 14. J Exp Bot. 2011. PMID: 21321051 Free PMC article.
-
Regulation of gene expression by jasmonate hormones.Phytochemistry. 2009 Sep;70(13-14):1560-70. doi: 10.1016/j.phytochem.2009.09.004. Epub 2009 Sep 30. Phytochemistry. 2009. PMID: 19796781 Review.
-
The plant Mediator complex and its role in jasmonate signaling.J Exp Bot. 2019 Jul 5;70(13):3415-3424. doi: 10.1093/jxb/erz233. J Exp Bot. 2019. PMID: 31089685 Free PMC article. Review.
Cited by
-
NaMYC2 transcription factor regulates a subset of plant defense responses in Nicotiana attenuata.BMC Plant Biol. 2013 May 1;13:73. doi: 10.1186/1471-2229-13-73. BMC Plant Biol. 2013. PMID: 23634896 Free PMC article.
-
The C2 Protein from the Geminivirus Tomato Yellow Leaf Curl Sardinia Virus Decreases Sensitivity to Jasmonates and Suppresses Jasmonate-Mediated Defences.Plants (Basel). 2016 Jan 15;5(1):8. doi: 10.3390/plants5010008. Plants (Basel). 2016. PMID: 27135228 Free PMC article.
-
Function and regulation of transcription factors involved in root apical meristem and stem cell maintenance.Front Plant Sci. 2015 Jul 6;6:505. doi: 10.3389/fpls.2015.00505. eCollection 2015. Front Plant Sci. 2015. PMID: 26217359 Free PMC article. Review.
-
Determinants of nucleosome positioning and their influence on plant gene expression.Genome Res. 2015 Aug;25(8):1182-95. doi: 10.1101/gr.188680.114. Epub 2015 Jun 10. Genome Res. 2015. PMID: 26063739 Free PMC article.
-
Regulation of Jasmonate-Induced Leaf Senescence by Antagonism between bHLH Subgroup IIIe and IIId Factors in Arabidopsis.Plant Cell. 2015 Jun;27(6):1634-49. doi: 10.1105/tpc.15.00110. Epub 2015 Jun 12. Plant Cell. 2015. PMID: 26071420 Free PMC article.
References
-
- Alonso J.M., Stepanova, A.N., Leisse, T.J., Kim, C.J., Chen, H., Shinn, P., Stevenson, D.K., Zimmerman, J., Barajas, P., Cheuk, R., et al. 2003. Genome wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653-657. - PubMed
-
- Amasino R.M. 1986. Acceleration of nucleic acid hybridization rate by polyethyleneglycol. Anal. Biochem. 152: 304-307. - PubMed
-
- Bechtold N. and Pelletier, G. 1998. In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Methods Mol. Biol. 82: 259-266. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials