

doi: 10.1128/JB.186.14.4829-4833.2004.
Reverse gyrase is not a prerequisite for hyperthermophilic life
Affiliations
- PMID: 15231817
- PMCID: PMC438624
- DOI: 10.1128/JB.186.14.4829-4833.2004
Item in Clipboard
Reverse gyrase is not a prerequisite for hyperthermophilic life
J Bacteriol.
2004 Jul.
Abstract
We disrupted the reverse gyrase gene from a hyperthermophilic archaeon, Thermococcus kodakaraensis KOD1. An apparent positive supercoiling activity that was observed in the host strain was not found in the disruptant strain. We found that a lack of reverse gyrase led to a retardation in growth that was more striking at higher temperatures. However, the disruption of the reverse gyrase gene did not lead to a lethal phenotype at 90 degrees C. This study provides experimental evidence that reverse gyrase is not a prerequisite for hyperthermophilic life.
Copyright 2004 American Society for Microbiology
Figures
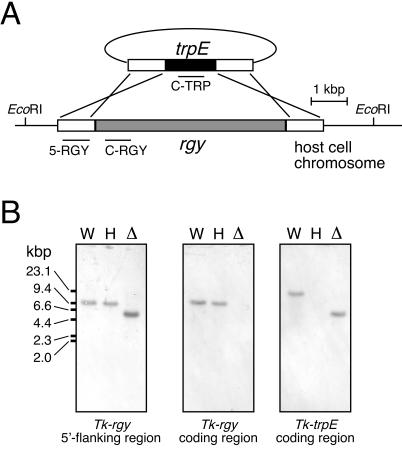
Disruption of rgyTk. (A) Strategy for disruption of the rgyTk gene via double crossover recombination. Regions corresponding to the probes used in the analysis described for panel B are indicated. (B) Confirmation of rgyTk gene disruption by Southern blot analyses. A digoxigenin-DNA labeling and detection kit (Roche Diagnostics) was used according to the manufacturer's instructions. The probes used were 5-RGY, a 728-bp fragment in the upstream region of the rgyTk gene (left panel), C-RGY, a 698-bp fragment within the rgyTk coding region (middle panel), and C-TRP, a 700-bp fragment within the T. kodakaraensis trpE coding region (right panel). Genome DNA from the wild type (W), the ΔtrpE host strain (H), and the Δrgy mutant strain (Δ) were digested with EcoRI. Results with 5-RGY show the decrease in length of the EcoRI fragment (from 9,360 to 5,641 bp) due to the replacement of the rgyTk gene with the T. kodakaraensis trpE gene, and those with C-RGY show the absence of the rgyTk gene on the Δrgy mutant strain chromosome.
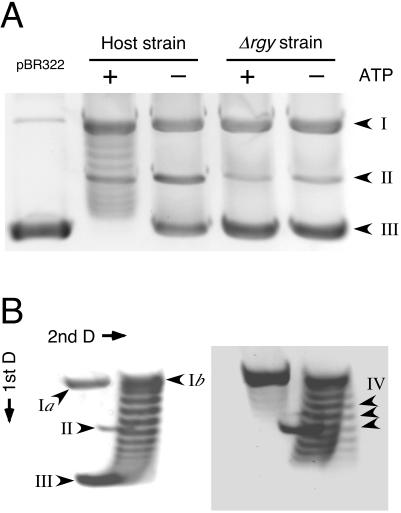
Comparison of DNA-relaxing activity and positive supercoiling activity in partially purified extracts of the host strain and the Δrgy strain. (A) DNA-relaxing activity in the partially purified extracts of the host strain and the Δrgy strain were analyzed as described in the text. The substrate was negatively supercoiled pBR322 (1 μg) purified from Escherichia coli (left lane). The presence (+) and absence (−) of ATP (2 mM) are indicated. (B) Positive supercoiling activity in the partially purified extracts of the host strain. Reaction conditions are described in the text, and ATP was added at a concentration of 2 mM. The second-dimension electrophoresis was performed in the presence of 4.5 μg of chloroquine per ml. Two stages of the reaction are shown. The left panel displays the progression of the reaction from negatively supercoiled DNA to relaxed DNA. The right panel displays the formation of positively supercoiled DNA. Different forms of pBR322 are indicated by the following arrowhead labels: I, nicked (Ia) or relaxed (Ib) DNA; II, linear DNA; III, negatively supercoiled DNA; IV, positively supercoiled DNA. D, dimension.
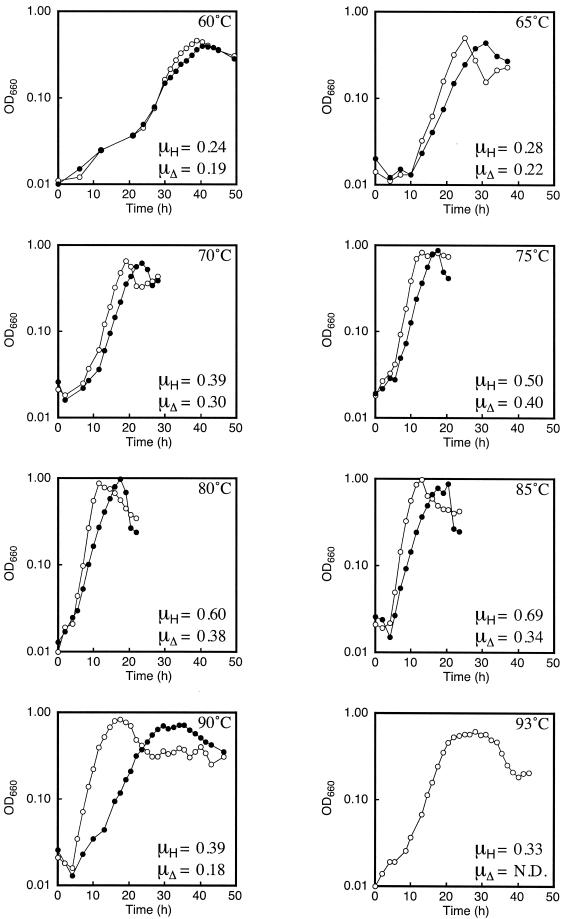
Growth curves and specific growth rates of the host strain and the Δrgy strain at various temperatures. Representative growth curves of the host strain and the Δrgy strain are shown. Temperatures are indicated in each panel. The specific growth rates of the host strain (μH) and those of the Δrgy strain (μΔ) were calculated from multiple growth curves. Open circles represent the host strain, and filled circles represent the Δrgy strain. N.D., not determined: OD660, optical density at 660 nm.
References
-
- Atomi, H., T. Fukui, T. Kanai, M. Morikawa, and T. Imanaka. 16 April 2004, posting date. Description of Thermococcus kodakaraensis sp. nov., a well studied hyperthermophilic archaeon previously reported as Pyrococcus sp. KOD1. Archaea [Online.] http://archaea.ws/archive/pdf/volume1/issue4/1-Imanaka.pdf. - PMC - PubMed
-
- Blöchl, E., R. Rachel, S. Burggraf, D. Hafenbradl, H. W. Jannasch, and K. O. Stetter. 1997. Pyrolobus fumarii, gen. and sp. nov., represents a novel group of archaea, extending the upper temperature limit for life to 113°C. Extremophiles 1:14-21. - PubMed
-
- Forterre, P. 2002. A hot story from comparative genomics: reverse gyrase is the only hyperthermophile-specific protein. Trends Genet. 18:236-237. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
