

BAD is a pro-survival factor prior to activation of its pro-apoptotic function
- PMID: 15231831
- PMCID: PMC4591964
- DOI: 10.1074/jbc.M406775200
BAD is a pro-survival factor prior to activation of its pro-apoptotic function
Abstract
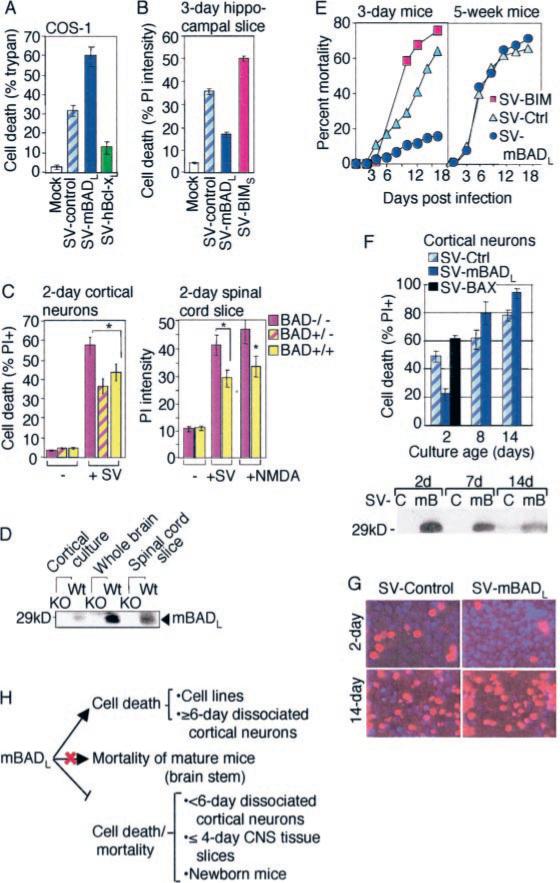
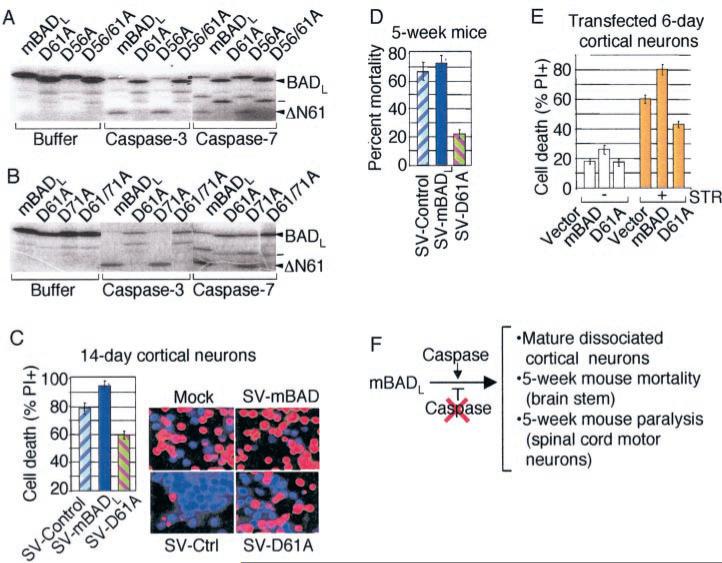
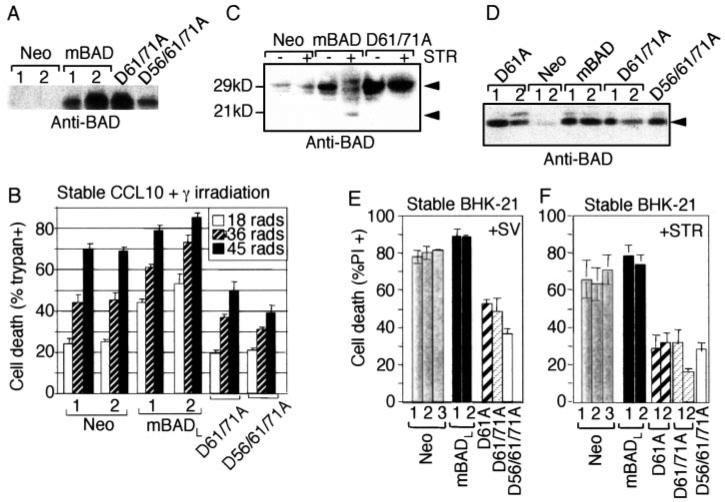
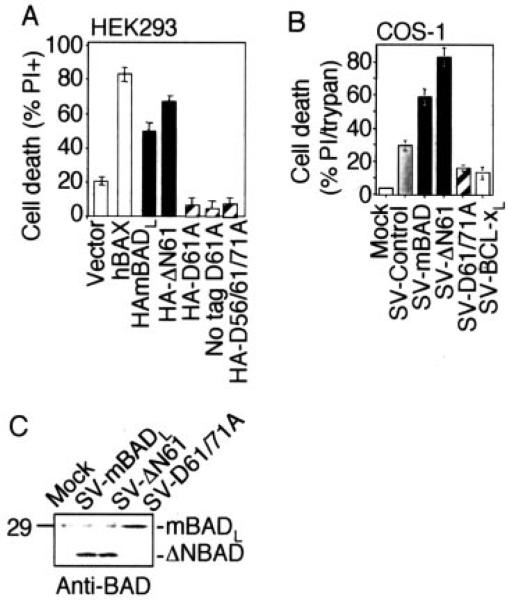
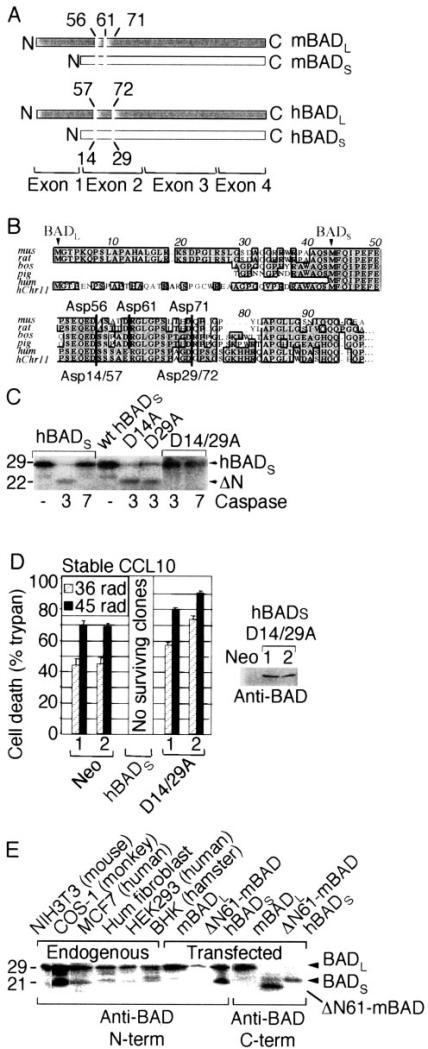
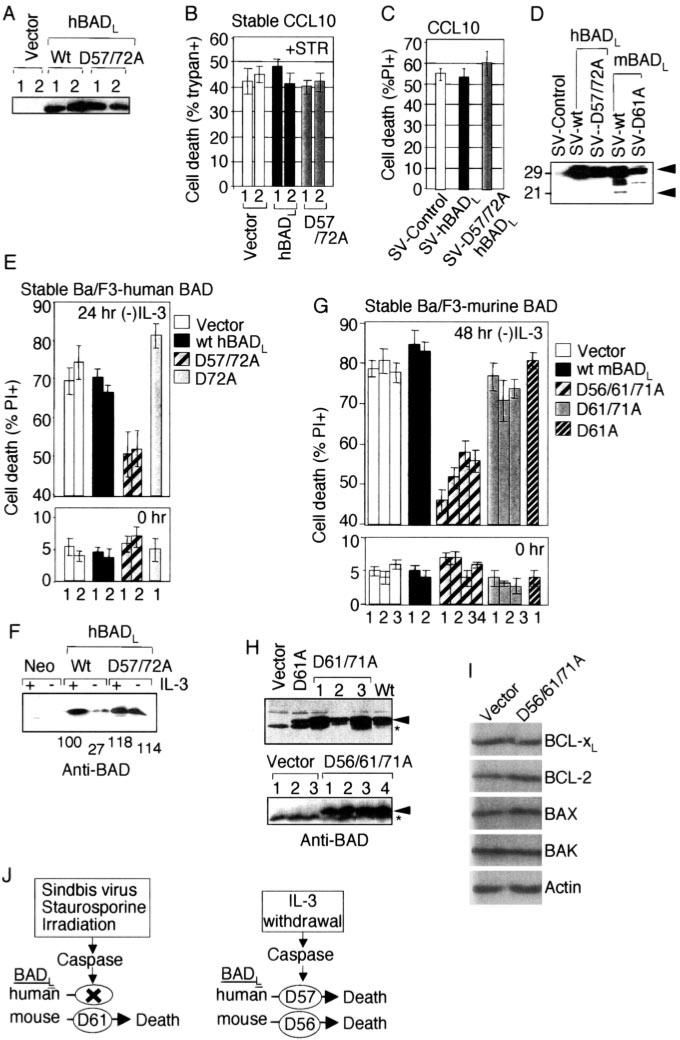
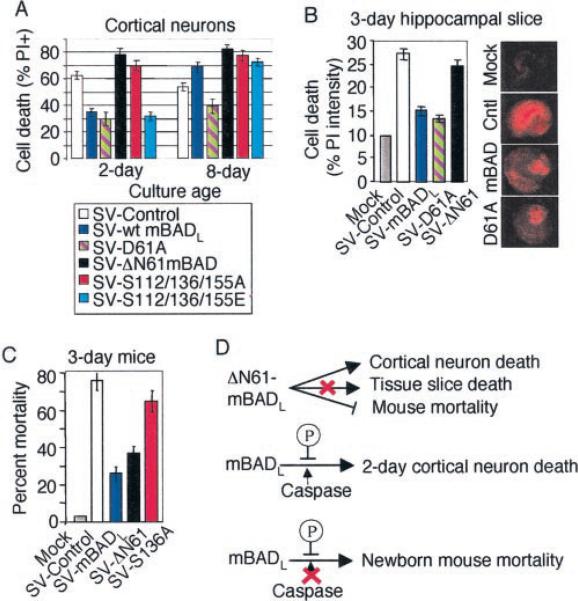
The mammalian BAD protein belongs to the BH3-only subgroup of the BCL-2 family. In contrast to its known pro-apoptotic function, we found that endogenous and overexpressed BAD(L) can inhibit cell death in neurons and other cell types. Several mechanisms regulate the conversion of BAD from an anti-death to a pro-death factor, including alternative splicing that produces the N-terminally truncated BAD(S). In addition, caspases convert BAD(L) into a pro-death fragment that resembles the short splice variant. The caspase site that is selectively cleaved during cell death following growth factor (interleukin-3) withdrawal is conserved between human and murine BAD. A second cleavage site that is required for murine BAD to promote death following Sindbis virus infection, gamma-irradiation, and staurosporine treatment is not conserved in human BAD, consistent with the inability of human BAD to promote death with these stimuli. However, loss of the BAD N terminus by any mechanism is not always sufficient to activate its pro-death activity, suggesting that the N terminus is a regulatory domain rather than an anti-death domain. These findings suggest that BAD is more than an inert death factor in healthy cells; it is also a pro-survival factor, prior to its role in promoting cell death.
Figures
Similar articles
-
Induction of Bad-mediated apoptosis by Sindbis virus infection: involvement of pro-survival members of the Bcl-2 family.Virology. 2002 Jan 20;292(2):258-71. doi: 10.1006/viro.2001.1206. Virology. 2002. PMID: 11878929
-
Caspase cleavage enhances the apoptosis-inducing effects of BAD.Mol Cell Biol. 2001 May;21(9):3025-36. doi: 10.1128/MCB.21.9.3025-3036.2001. Mol Cell Biol. 2001. PMID: 11287608 Free PMC article.
-
A comparison of the properties of a Bcl-xL variant to the wild-type anti-apoptosis inhibitor in mammalian cell cultures.Metab Eng. 2003 Oct;5(4):230-45. doi: 10.1016/s1096-7176(03)00044-2. Metab Eng. 2003. PMID: 14642351
-
Structural biology of the Bcl-2 family of proteins.Biochim Biophys Acta. 2004 Mar 1;1644(2-3):83-94. doi: 10.1016/j.bbamcr.2003.08.012. Biochim Biophys Acta. 2004. PMID: 14996493 Review.
-
Death and survival signals determine active/inactive conformations of pro-apoptotic BAX, BAD, and BID molecules.Cold Spring Harb Symp Quant Biol. 1999;64:343-50. doi: 10.1101/sqb.1999.64.343. Cold Spring Harb Symp Quant Biol. 1999. PMID: 11232306 Review. No abstract available.
Cited by
-
Influence of stochastic gene expression on the cell survival rheostat after traumatic brain injury.PLoS One. 2011;6(8):e23111. doi: 10.1371/journal.pone.0023111. Epub 2011 Aug 11. PLoS One. 2011. PMID: 21853077 Free PMC article.
-
bcl2, bax, and nestin in the brains of patients with neurodegeneration and those of normal aging.J Mol Neurosci. 2005;27(2):167-74. doi: 10.1385/JMN:27:2:167. J Mol Neurosci. 2005. PMID: 16186627
-
Influenza virus induces apoptosis via BAD-mediated mitochondrial dysregulation.J Virol. 2013 Jan;87(2):1049-60. doi: 10.1128/JVI.02017-12. Epub 2012 Nov 7. J Virol. 2013. PMID: 23135712 Free PMC article.
-
Multiple functions of BCL-2 family proteins.Cold Spring Harb Perspect Biol. 2013 Feb 1;5(2):a008722. doi: 10.1101/cshperspect.a008722. Cold Spring Harb Perspect Biol. 2013. PMID: 23378584 Free PMC article. Review.
-
Multipolar functions of BCL-2 proteins link energetics to apoptosis.Trends Cell Biol. 2012 Jun;22(6):318-28. doi: 10.1016/j.tcb.2012.03.005. Epub 2012 May 5. Trends Cell Biol. 2012. PMID: 22560661 Free PMC article. Review.
References
-
- Lewis J, Oyler GA, Ueno K, Fannjiang Y, Chau BN, Vornov J, Korsmeyer SJ, Zou S, Hardwick JM. Nat. Med. 1999;5:832–835. - PubMed
-
- Middleton G, Davies AM. Development (Camb.) 2001;128:4715–4728. - PubMed
-
- Fannjiang Y, Kim C-H, Huganir RL, Zou RL, S., Lindsten T, Thompson CB, Mito T, Traystman RJ, Larsen T, Griffin DE, Mandir AS, Dawson TM, Dike S, Sappington AL, Kerr DA, Jonas EA, Kaczmarek LK, Hardwick JM. Dev. Cell. 2003;4:575–585. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
