

Extracellular calcium acts as a "third messenger" to regulate enzyme and alkaline secretion
- PMID: 15240573
- PMCID: PMC2172151
- DOI: 10.1083/jcb.200310145
Extracellular calcium acts as a "third messenger" to regulate enzyme and alkaline secretion
Abstract
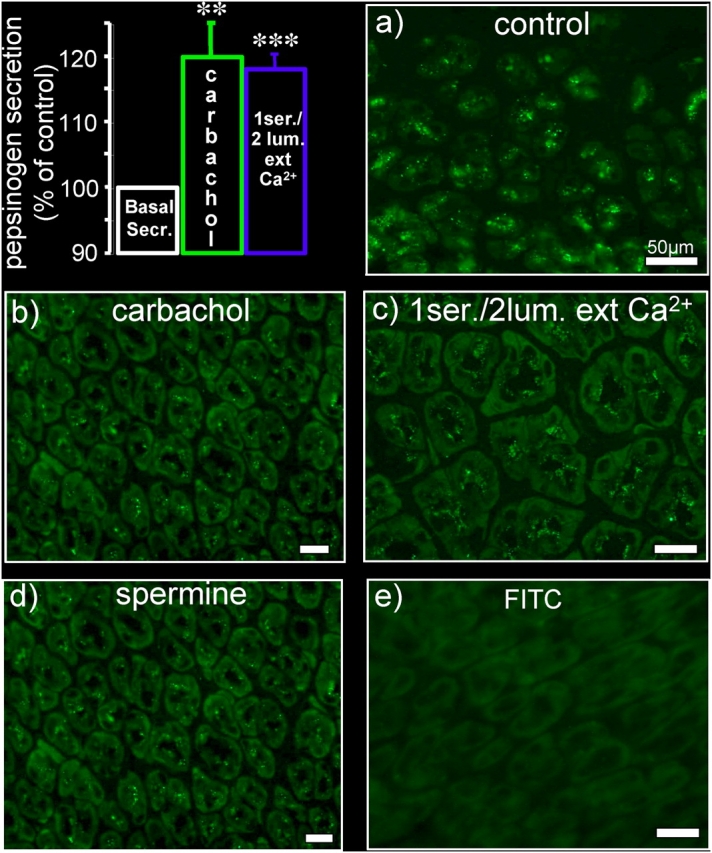
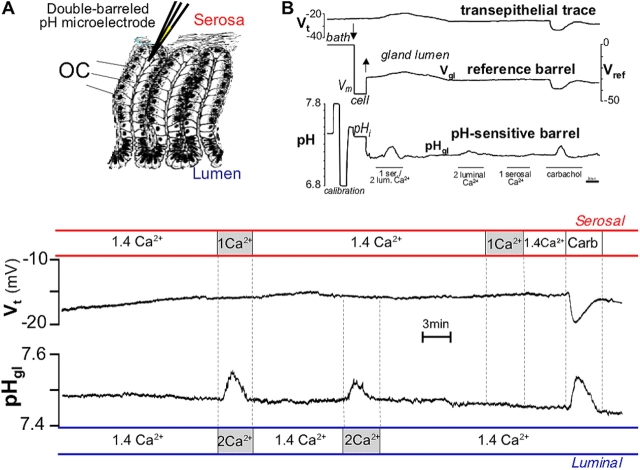
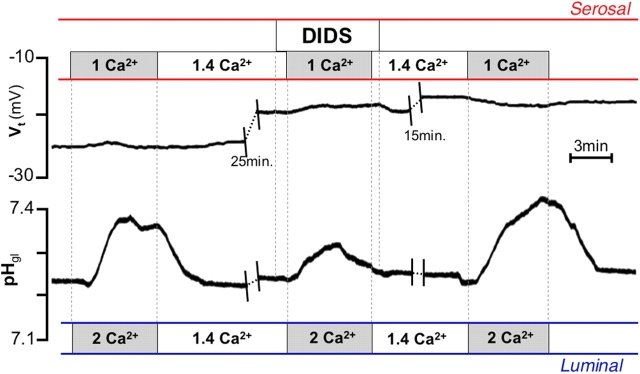
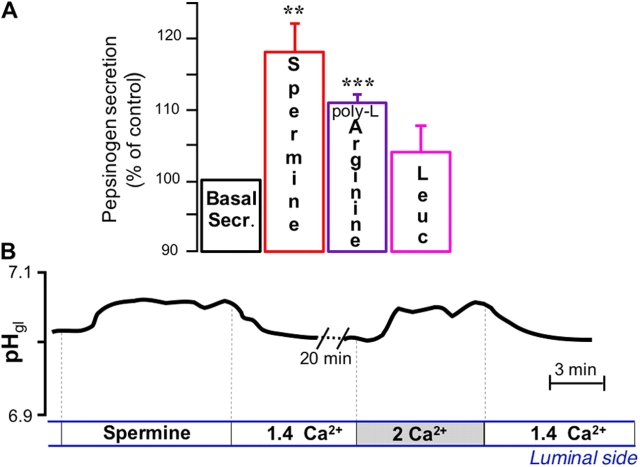
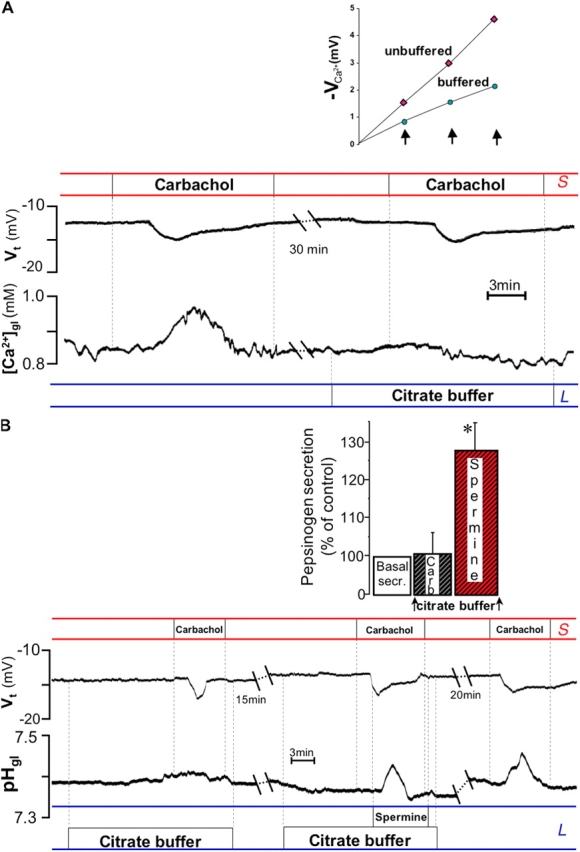
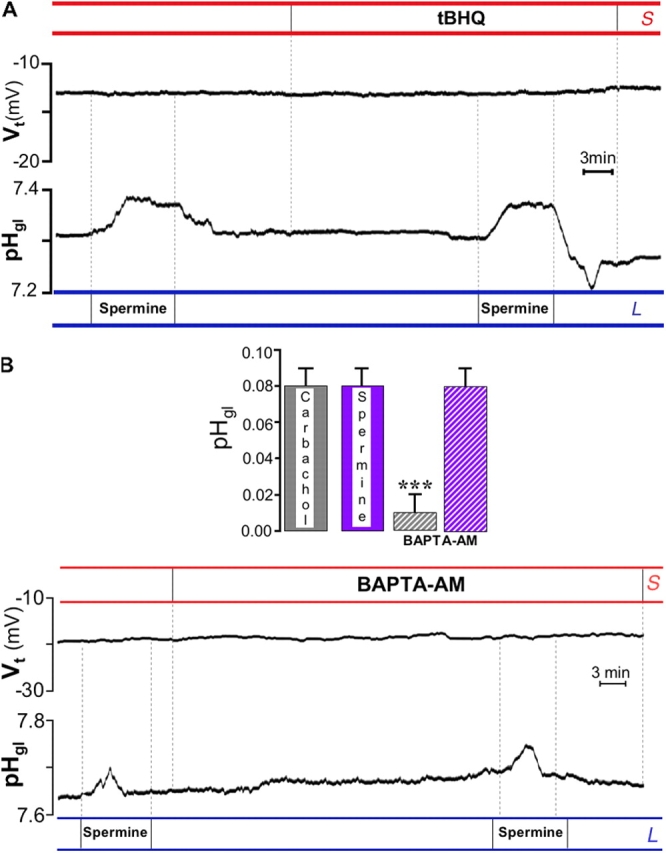
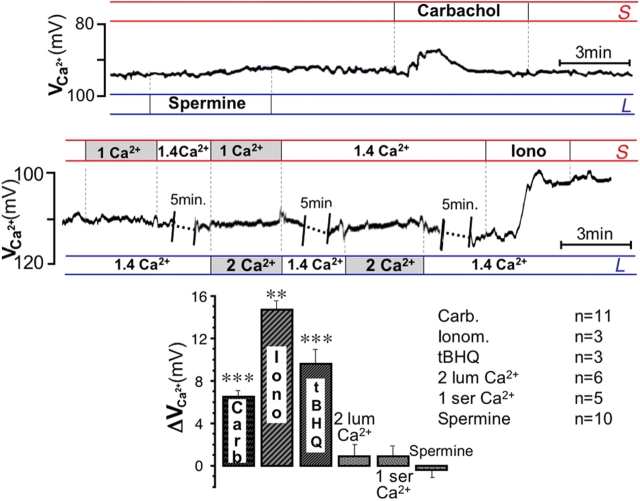
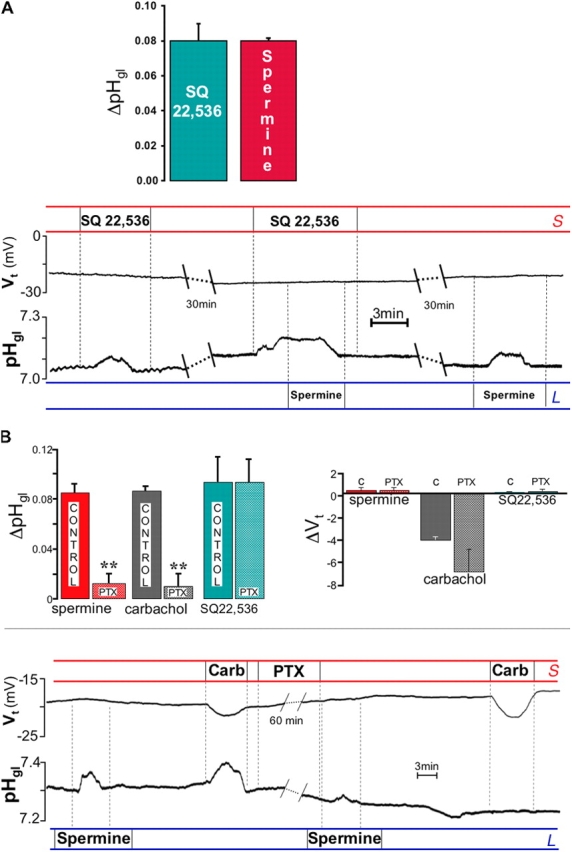
It is generally assumed that the functional consequences of stimulation with Ca2+ -mobilizing agonists are derived exclusively from the second messenger action of intracellular Ca2+, acting on targets inside the cells. However, during Ca2+ signaling events, Ca2+ moves in and out of the cell, causing changes not only in intracellular Ca2+, but also in local extracellular Ca2+. The fact that numerous cell types possess an extracellular Ca2+ "sensor" raises the question of whether these dynamic changes in external [Ca2+] may serve some sort of messenger function. We found that in intact gastric mucosa, the changes in extracellular [Ca2+] secondary to carbachol-induced increases in intracellular [Ca2+] were sufficient and necessary to elicit alkaline secretion and pepsinogen secretion, independent of intracellular [Ca2+] changes. These findings suggest that extracellular Ca2+ can act as a "third messenger" via Ca2+ sensor(s) to regulate specific subsets of tissue function previously assumed to be under the direct control of intracellular Ca2+.
Figures
Similar articles
-
Cellular mechanisms of carbachol-stimulated Cl- secretion in rat epididymal epithelium.Biol Reprod. 2006 Sep;75(3):407-13. doi: 10.1095/biolreprod.106.052316. Epub 2006 Jun 7. Biol Reprod. 2006. PMID: 16760378
-
The effect of sevoflurane on intracellular calcium concentration from cholinergic cells.Brain Res Bull. 2006 Mar 31;69(2):147-52. doi: 10.1016/j.brainresbull.2005.11.016. Epub 2005 Dec 19. Brain Res Bull. 2006. PMID: 16533663
-
Cholinergic desensitization of pepsinogen secretion and calcium mobilization of dispersed guinea pig chief cells.J Cell Physiol. 1990 Jan;142(1):46-54. doi: 10.1002/jcp.1041420107. J Cell Physiol. 1990. PMID: 2298823
-
Modulation of muscarinic signaling in PC12 cells overexpressing neuronal Ca2+ sensor-1 protein.Cell Mol Biol (Noisy-le-grand). 2009 Aug 5;55 Suppl:OL1138-50. Cell Mol Biol (Noisy-le-grand). 2009. PMID: 19656467
-
The role of Ca2+ influx and intracellular Ca2+ release in the muscarinic-mediated contraction of mammalian urinary bladder smooth muscle.BJU Int. 2006 Oct;98(4):868-75. doi: 10.1111/j.1464-410X.2006.06431.x. BJU Int. 2006. PMID: 16978287
Cited by
-
In vivo epithelial wound repair requires mobilization of endogenous intracellular and extracellular calcium.J Biol Chem. 2013 Nov 22;288(47):33585-33597. doi: 10.1074/jbc.M113.488098. Epub 2013 Oct 11. J Biol Chem. 2013. PMID: 24121509 Free PMC article.
-
Termination of cAMP signals by Ca2+ and G(alpha)i via extracellular Ca2+ sensors: a link to intracellular Ca2+ oscillations.J Cell Biol. 2005 Oct 24;171(2):303-12. doi: 10.1083/jcb.200507054. J Cell Biol. 2005. PMID: 16247029 Free PMC article.
-
The Different Facets of Extracellular Calcium Sensors: Old and New Concepts in Calcium-Sensing Receptor Signalling and Pharmacology.Int J Mol Sci. 2018 Mar 27;19(4):999. doi: 10.3390/ijms19040999. Int J Mol Sci. 2018. PMID: 29584660 Free PMC article. Review.
-
Importance of Ca(2+) in gastric epithelial restitution-new views revealed by real-time in vivo measurements.Curr Opin Pharmacol. 2014 Dec;19:76-83. doi: 10.1016/j.coph.2014.07.012. Epub 2014 Aug 9. Curr Opin Pharmacol. 2014. PMID: 25108560 Free PMC article. Review.
-
Calcium as an extracellular signalling molecule: perspectives on the Calcium Sensing Receptor in the brain.C R Biol. 2005 Aug;328(8):691-700. doi: 10.1016/j.crvi.2004.10.006. Epub 2005 Feb 8. C R Biol. 2005. PMID: 16125647 Free PMC article. Review.
References
-
- Ashby, M.C., and A.V. Tepikin. 2002. Polarized calcium and calmodulin signaling in secretory epithelia. Physiol. Rev. 82:701–734. - PubMed
-
- Babini, E., M. Paukert, H.S. Geisler, and S. Grunder. 2002. Alternative splicing and interaction with di- and polyvalent cations control the dynamic range of acid-sensing ion channel 1 (ASIC1). J. Biol. Chem. 277:41597–41603. - PubMed
-
- Belan, P., O. Gerasimenko, A. Tepikin, and O. Petersen. 1996. Localization of Ca2+ extrusion sites in pancreatic acinar cells. J. Biol. Chem. 271:7615–7619. - PubMed
-
- Belan, P., O. Gerasimenko, O.H. Petersen, and A.V. Tepikin. 1997. Distribution of Ca2+ extrusion sites on the mouse pancreatic acinar cell surface. Cell Calcium. 22:5–10. - PubMed
-
- Berridge, M.J., M.D. Bootman, and H.L. Roderick. 2003. Calcium signalling: dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 4:517–529. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
