

Specific engagement of TLR4 or TLR3 does not lead to IFN-beta-mediated innate signal amplification and STAT1 phosphorylation in resident murine alveolar macrophages
- PMID: 15240691
- PMCID: PMC2701405
- DOI: 10.4049/jimmunol.173.2.1033
Specific engagement of TLR4 or TLR3 does not lead to IFN-beta-mediated innate signal amplification and STAT1 phosphorylation in resident murine alveolar macrophages
Abstract
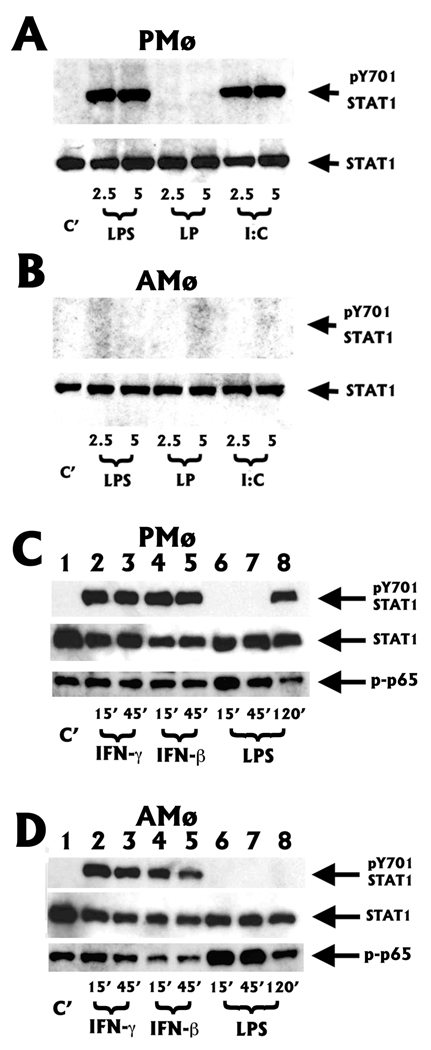
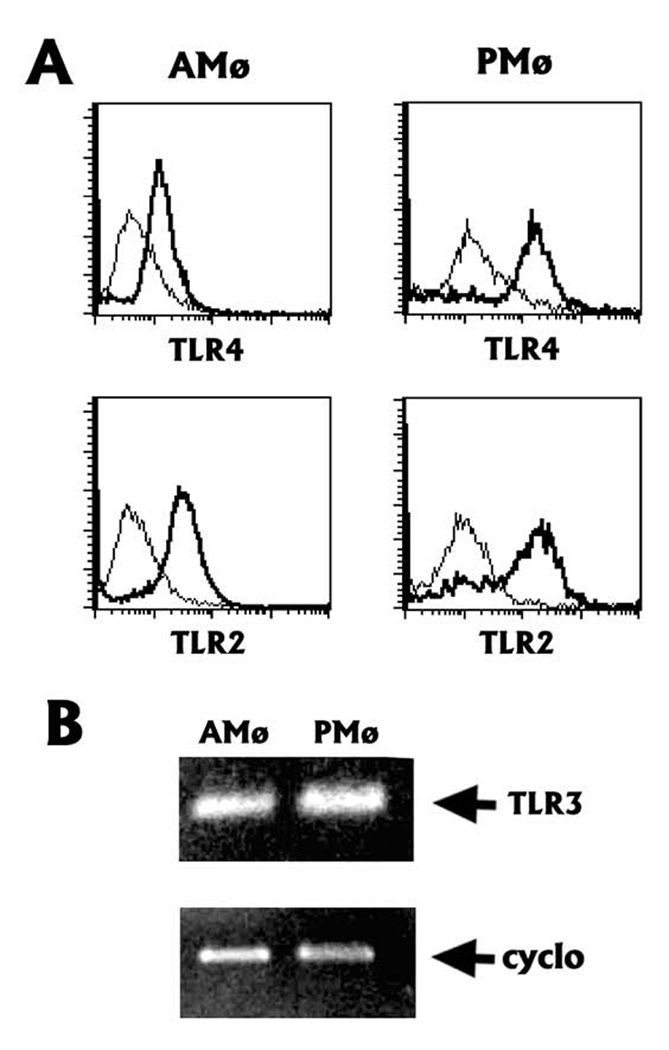
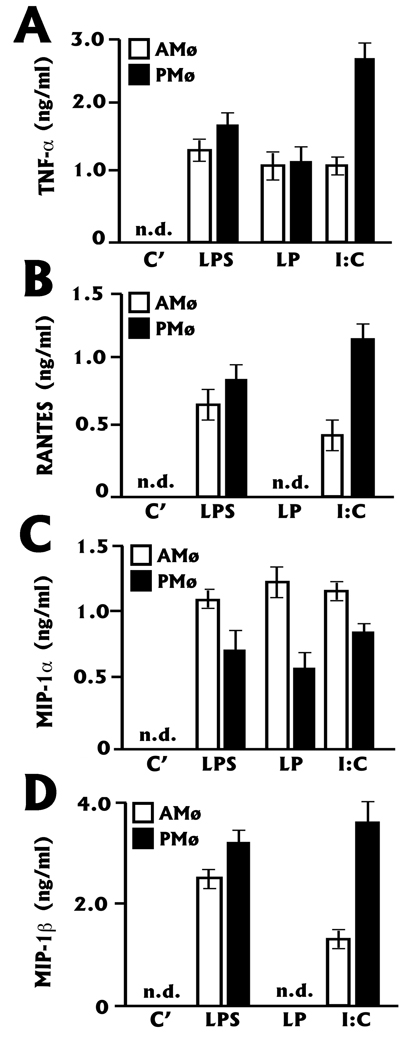
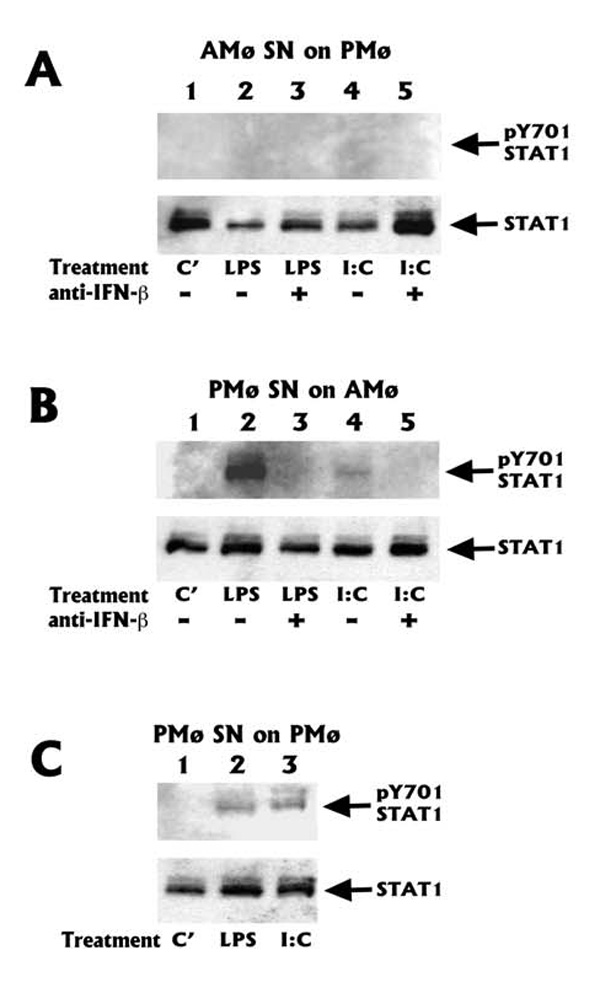
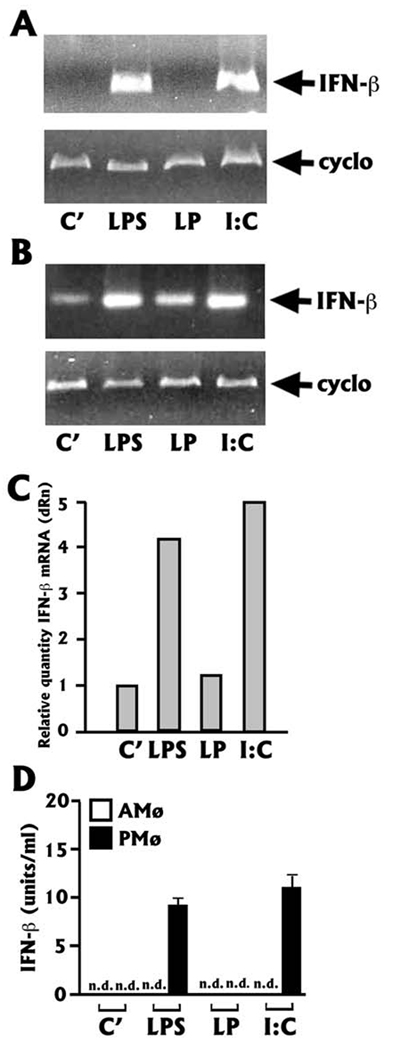
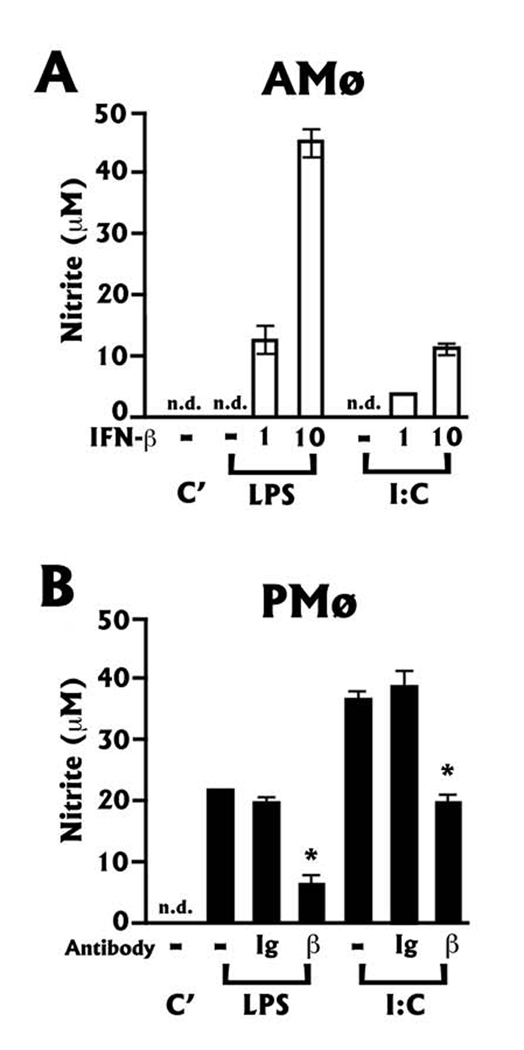
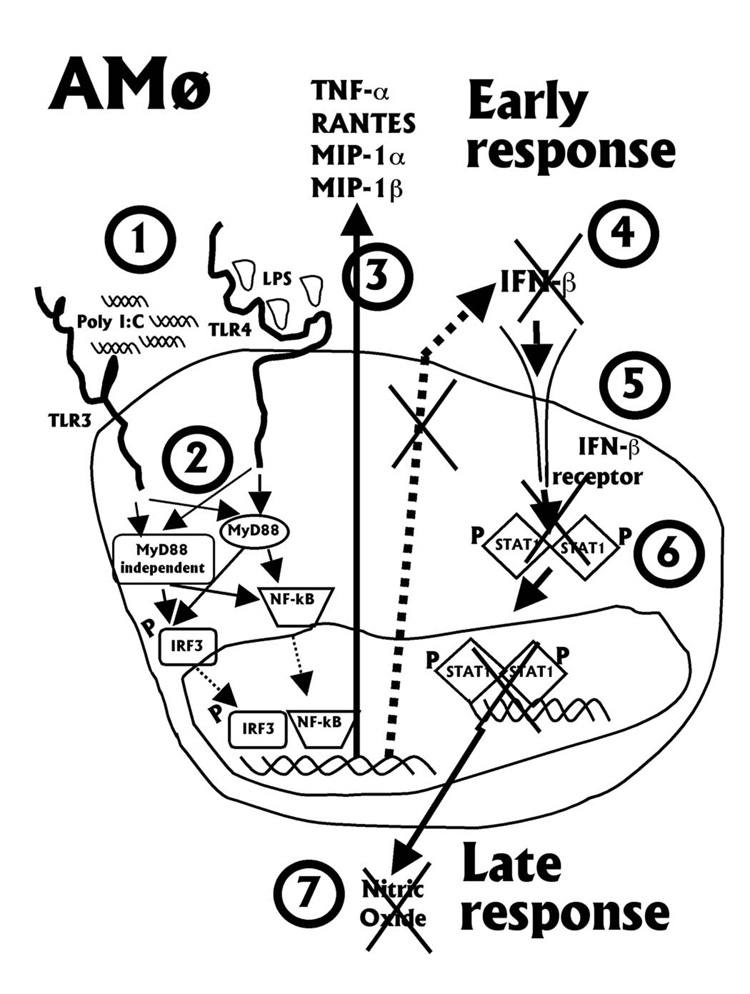
The innate immune response must be mobilized promptly yet judiciously via TLRs to protect the lungs against pathogens. Stimulation of murine peritoneal macrophage (PMphi) TLR4 or TLR3 by pathogen-associated molecular patterns (PAMPs) typically induces type I IFN-beta, leading to autocrine activation of the transcription factor STAT1. Because it is unknown whether STAT1 plays a similar role in the lungs, we studied the response of resident alveolar macrophages (AMphi) or control PMphi from normal C57BL/6 mice to stimulation by PAMPs derived from viruses (polyriboinosinic:polyribocytidylic acid, specific for TLR3) or bacteria (Pam(3)Cys, specific for TLR2, and repurified LPS, specific for TLR4). AMphi did not activate STAT1 by tyrosine phosphorylation on Y701 following stimulation of any of these three TLRs, but readily did so in response to exogenous IFN-beta. This unique AMphi response was not due to altered TLR expression, or defective immediate-early gene response, as measured by expression of TNF-alpha and three beta chemokines. Instead, AMphi differed from PMphi in not producing bioactive IFN-beta, as confirmed by ELISA and by the failure of supernatants from TLR-stimulated AMphi to induce STAT1 phosphorylation in PMphi. Consequently, AMphi did not produce the microbicidal effector molecule NO following TLR4 or TLR3 stimulation unless exogenous IFN-beta was also added. Thus, murine AMphi respond to bacterial or viral PAMPs by producing inflammatory cytokines and chemokines, but because they lack the feed-forward amplification typically mediated by autocrine IFN-beta secretion and STAT1 activation, require exogenous IFN to mount a second phase of host defense.
Figures
Similar articles
-
Toll-like receptor 3 mediates a more potent antiviral response than Toll-like receptor 4.J Immunol. 2003 Apr 1;170(7):3565-71. doi: 10.4049/jimmunol.170.7.3565. J Immunol. 2003. PMID: 12646618
-
TLR4, but not TLR2, mediates IFN-beta-induced STAT1alpha/beta-dependent gene expression in macrophages.Nat Immunol. 2002 Apr;3(4):392-8. doi: 10.1038/ni774. Epub 2002 Mar 18. Nat Immunol. 2002. PMID: 11896392
-
TLR2 and TLR4 agonists stimulate unique repertoires of host resistance genes in murine macrophages: interferon-beta-dependent signaling in TLR4-mediated responses.J Endotoxin Res. 2003;9(3):169-75. doi: 10.1179/096805103125001577. J Endotoxin Res. 2003. PMID: 12831458
-
Toll-like receptors as adjuvant receptors.Biochim Biophys Acta. 2002 Feb 13;1589(1):1-13. doi: 10.1016/s0167-4889(01)00182-3. Biochim Biophys Acta. 2002. PMID: 11909637 Review.
-
Microbial recognition by Toll-like receptors.J Dermatol Sci. 2004 Apr;34(2):73-82. doi: 10.1016/j.jdermsci.2003.10.002. J Dermatol Sci. 2004. PMID: 15033189 Review.
Cited by
-
Soybean-derived Bowman-Birk Inhibitor (BBI) Inhibits HIV Replication in Macrophages.Sci Rep. 2016 Oct 13;6:34752. doi: 10.1038/srep34752. Sci Rep. 2016. PMID: 27734899 Free PMC article.
-
Differential type I interferon activation and susceptibility of dendritic cell populations to porcine arterivirus.Immunology. 2007 Feb;120(2):217-29. doi: 10.1111/j.1365-2567.2006.02493.x. Epub 2006 Nov 20. Immunology. 2007. PMID: 17116172 Free PMC article.
-
Cell-mediated adaptive immune defense of the lungs.Proc Am Thorac Soc. 2005;2(5):412-6. doi: 10.1513/pats.200507-070JS. Proc Am Thorac Soc. 2005. PMID: 16322591 Free PMC article. Review.
-
Compartmentalization of the inflammatory response during bacterial sepsis and severe COVID-19.J Intensive Med. 2024 Feb 27;4(3):326-340. doi: 10.1016/j.jointm.2024.01.001. eCollection 2024 Jul. J Intensive Med. 2024. PMID: 39035623 Free PMC article. Review.
-
A Pathophysiological Insight into Sepsis and Its Correlation with Postmortem Diagnosis.Mediators Inflamm. 2016;2016:4062829. doi: 10.1155/2016/4062829. Epub 2016 Apr 27. Mediators Inflamm. 2016. PMID: 27239102 Free PMC article. Review.
References
-
- Barnes PJ. New concepts in chronic obstructive pulmonary disease. Annu Rev Med. 2003;54:113. - PubMed
-
- Tetley TD. Macrophages and the pathogenesis of COPD. Chest. 2002;121:156S. - PubMed
-
- Hodge S, Hodge G, Scicchitano R, Reynolds PN, Holmes M. Alveolar macrophages from subjects with chronic obstructive pulmonary disease are deficient in their ability to phagocytose apoptotic airway epithelial cells. Immunol Cell Biol. 2003;81:289. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
