

Rapid axoglial signaling mediated by neuregulin and neurotrophic factors
- PMID: 15240814
- PMCID: PMC6729661
- DOI: 10.1523/JNEUROSCI.1692-04.2004
Rapid axoglial signaling mediated by neuregulin and neurotrophic factors
Erratum in
- J Neurosci. 2004 Jul 28;24(30):1 p following 6852
Abstract
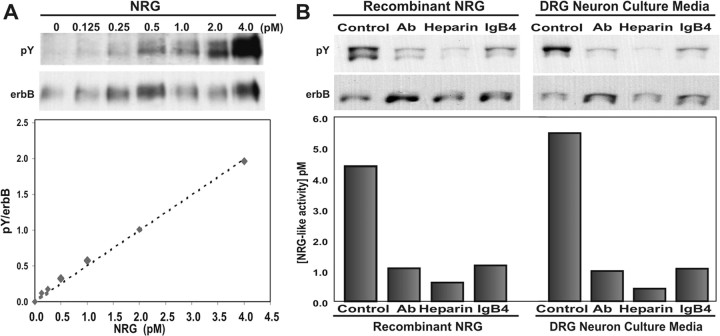
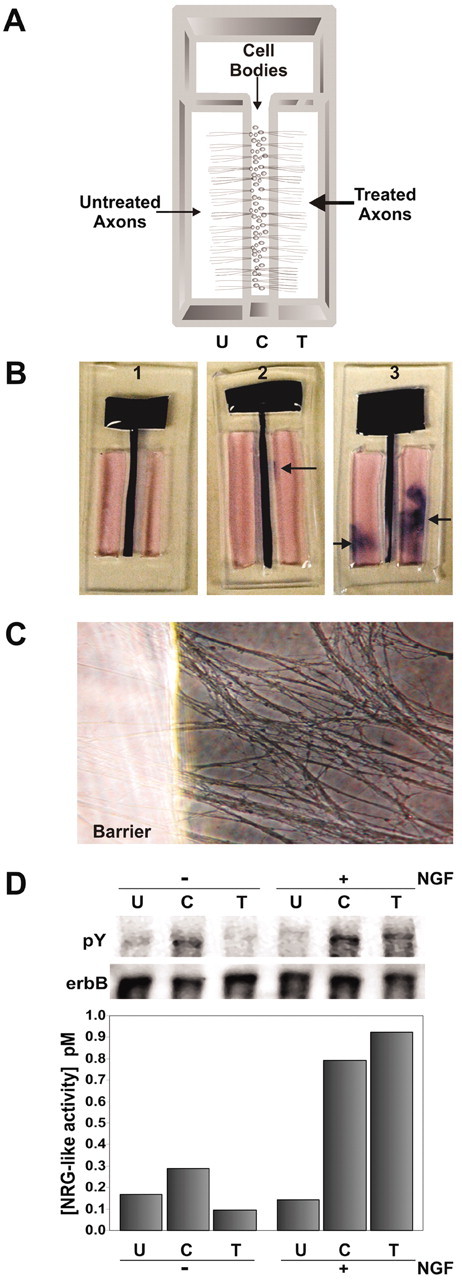
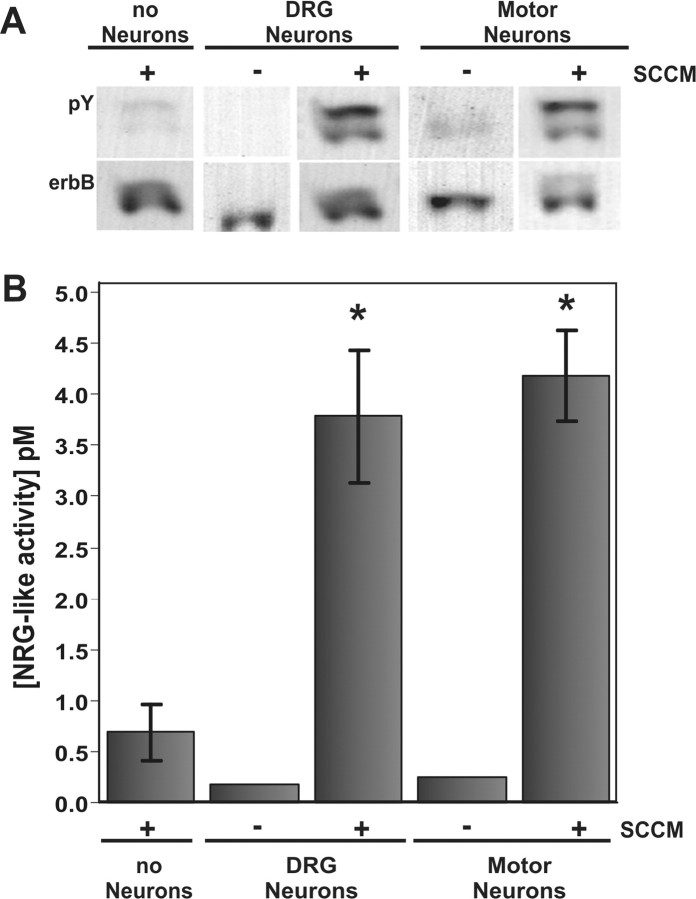
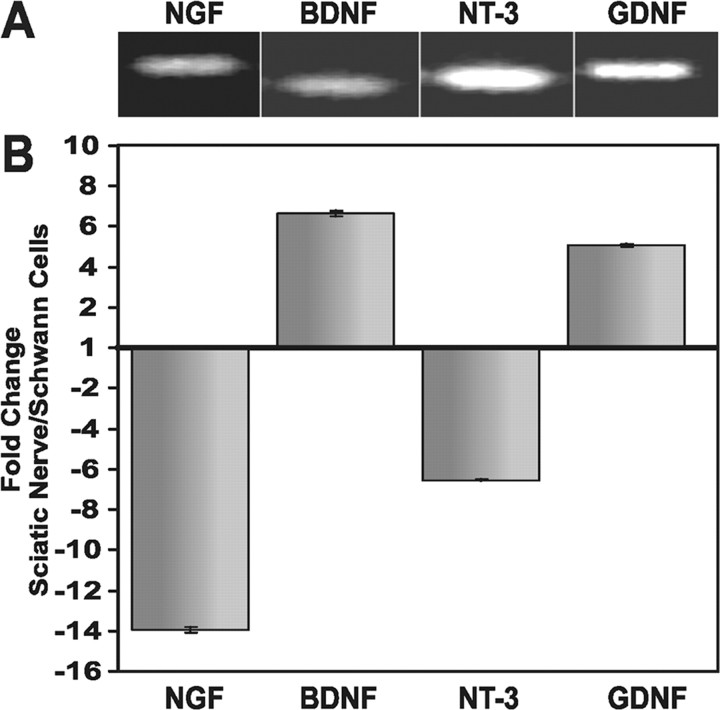
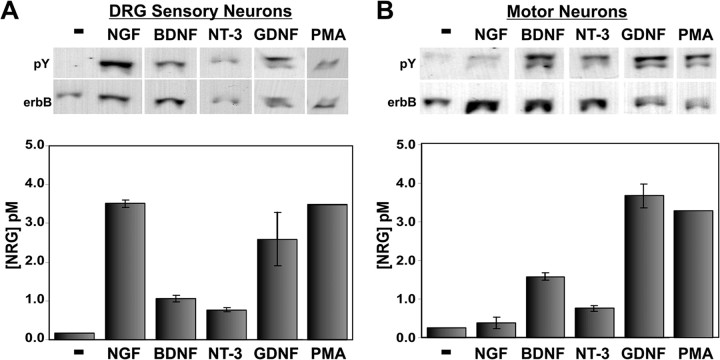
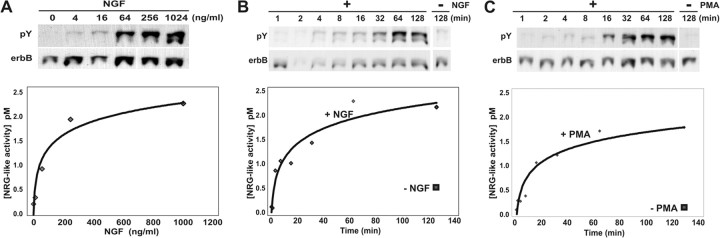
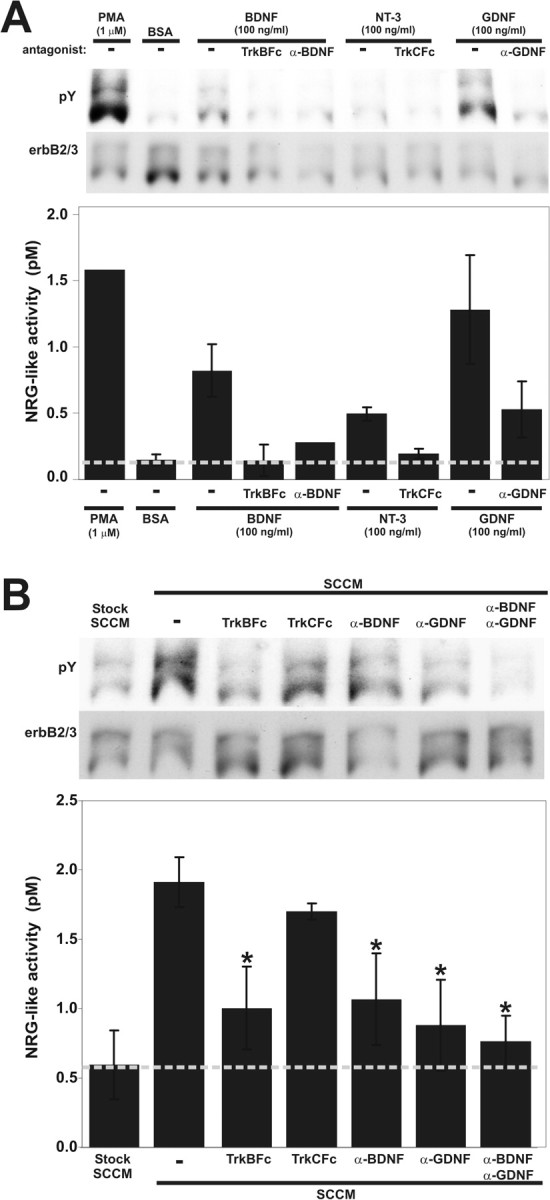
During peripheral nervous system development, Schwann cells are precisely matched to the axons that they support. This is mediated by axonal neuregulins that are essential for Schwann cell survival and differentiation. Here, we show that sensory and motor axons rapidly release heparin-binding forms of neuregulin in response to Schwann cell-derived neurotrophic factors in a dose-dependent manner. Neuregulin release occurs within minutes, is saturable, and occurs from axons that were isolated using a newly designed chamber slide apparatus. Although NGF and glial cell line-derived neurotrophic factor (GDNF) were the most potent neurotrophic factors to release neuregulin from sensory neurons, GDNF and BDNF were most potent for motor neurons and were the predominant neuregulin-releasing neurotrophic factors produced by cultured Schwann cells. Comparable levels of neuregulin could be released at a similar rate from neurons after protein kinase C activation with the phorbol ester, phorbol 12-myristate 13-acetate, which has also been shown to promote the cleavage and release of neuregulin from its transmembrane precursor. The rapid release of neuregulin from axons in response to Schwann cell-derived neurotrophic factors may be part of a spatially restricted system of communication at the axoglial interface important for proper peripheral nerve development, function, and repair.
Figures
References
-
- Adlkofer K, Lai C (2000) Role of neuregulins in glial cell development. Glia 29: 104-111. - PubMed
-
- Apfel SC (1999) Neurotrophic factors and diabetic peripheral neuropathy. Eur Neurol 41 [Suppl 1]: 27-34. - PubMed
-
- Baek SY, Kim SU (1998) Proliferation of human Schwann cells induced by neu differentiation factor isoforms. Dev Neurosci 20: 512-517. - PubMed
-
- Baig MA, Khan MA (1996) The induction of neurotrophin and TRK receptor mRNA expression during early avian embryogenesis. Int J Dev Neurosci 14: 55-60. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources