

Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion, and community assembly
- PMID: 15243158
- PMCID: PMC503710
- DOI: 10.1073/pnas.0403458101
Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion, and community assembly
Abstract
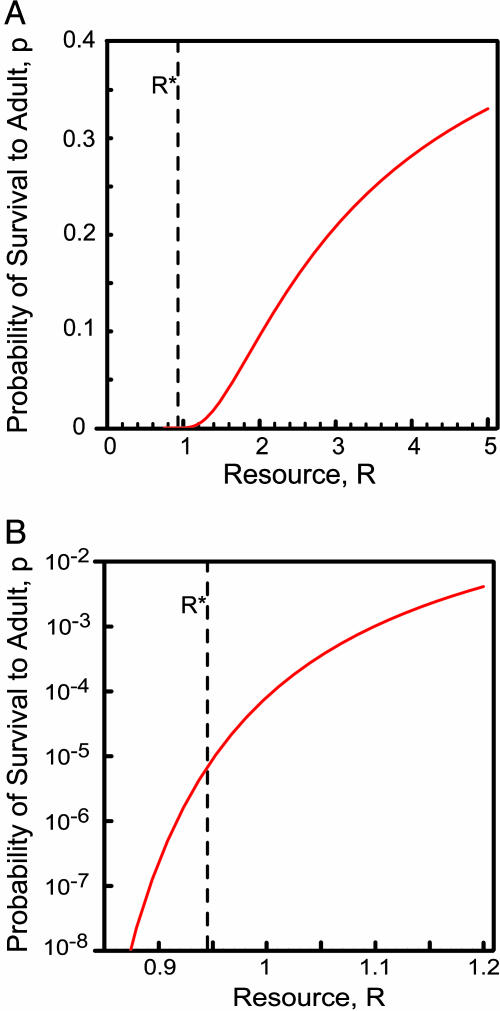
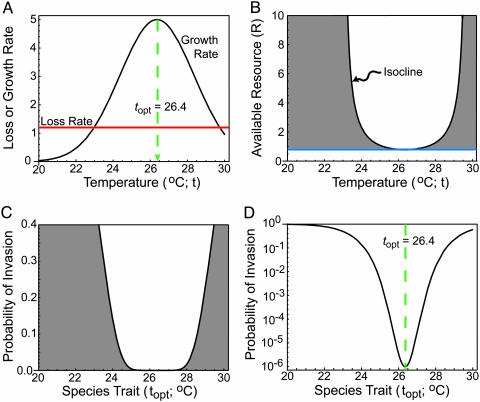
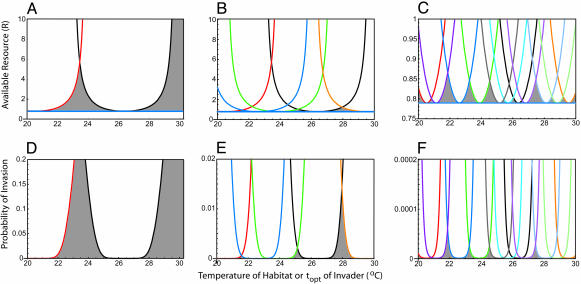
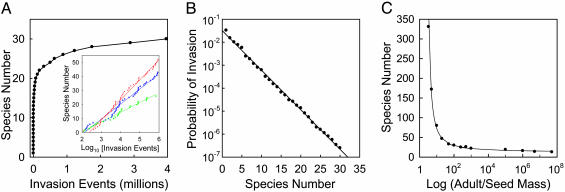
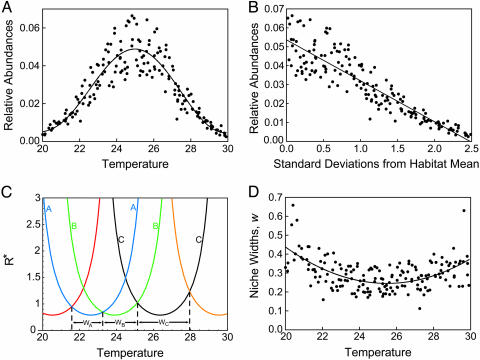
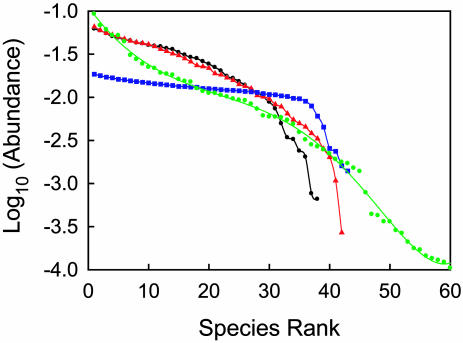
Stochastic niche theory resolves many of the differences between neutral theory and classical tradeoff-based niche theories of resource competition and community structure. In stochastic niche theory, invading species become established only if propagules can survive stochastic mortality while growing to maturity on the resources left unconsumed by established species. The theory makes three predictions about community structure. First, stochastic niche assembly creates communities in which species dominate approximately equally wide "slices" of the habitat's spatial heterogeneity. These niche widths generate realistic distributions of species relative abundances for which, contrary to neutral theory but consistent with numerous observations, there are strong correlations among species traits, species abundances, and environmental conditions. Second, slight decreases in resource levels are predicted to cause large decreases in the probability that a propagule would survive to be an adult. These decreases cause local diversity to be limited by the inhibitory effects of resource use by established species on the establishment (recruitment) of potential invaders. If resource pulses or disturbance allowed invaders to overcome this recruitment limitation, many more species could indefinitely coexist. Third, the low invasibility of high diversity communities is predicted to result not from diversity per se, but from the uniformly low levels of resources that occur in high-diversity communities created by stochastic competitive assembly. This prediction provides a potential solution to the invasion paradox, which is the tendency for highly diverse regions to be more heavily invaded.
Figures
References
-
- Hastings, A. (1980) Theor. Popul. Biol. 18, 363–373. - PubMed
-
- Levins, R. (1979) Am. Nat. 114, 765–783.
-
- Tilman, D. (1976) Science 192, 463–465. - PubMed
-
- Tilman, D. (1982) Resource Competition and Community Structure, Monographs in Population Biology (Princeton Univ. Press, Princeton). - PubMed
-
- Tilman, D. (1988) Plant Strategies and the Dynamics and Structure of Plant Communities (Princeton Univ. Press, Princeton).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
