

Anti-apoptotic protein Bcl-2 interacts with and destabilizes the sarcoplasmic/endoplasmic reticulum Ca2+-ATPase (SERCA)
- PMID: 15245329
- PMCID: PMC1134078
- DOI: 10.1042/BJ20040187
Anti-apoptotic protein Bcl-2 interacts with and destabilizes the sarcoplasmic/endoplasmic reticulum Ca2+-ATPase (SERCA)
Abstract
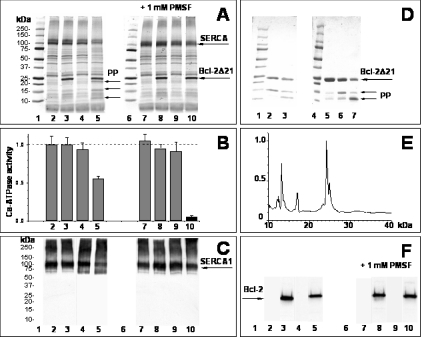
The anti-apoptotic effect of Bcl-2 is well established, but the detailed mechanisms are unknown. In the present study, we show in vitro a direct interaction of Bcl-2 with the rat skeletal muscle SERCA (sarcoplasmic/endoplasmic reticulum Ca2+-ATPase), leading to destabilization and inactivation of the protein. Recombinant human Bcl-2D21, a truncated form of Bcl-2 with a deletion of 21 residues at the C-terminal membrane-anchoring region, was expressed and affinity-purified as a glutathione S-transferase fusion protein. Bcl-2D21 co-immunoprecipitated and specifically interacted with SERCA in an in vitro-binding assay. The original level of Bcl-2 in sarcoplasmic reticulum vesicles was very low, i.e. hardly detectable by immunoblotting with specific antibodies. The addition of Bcl-2D21 to the sarcoplasmic reticulum resulted in the inhibition of the Ca2+-ATPase activity dependent on the Bcl-2D21/SERCA molar ratio and incubation time. A complete inactivation of SERCA was observed after 2.5 h of incubation at approx. 2:1 molar ratio of Bcl-2D21 to SERCA. In contrast, Bcl-2D21 did not significantly change the activity of the plasma-membrane Ca2+-ATPase. The redox state of the single Cys158 residue in Bcl-2D21 and the presence of GSH did not affect SERCA inhibition. The interaction of Bcl-2D21 with SERCA resulted in a conformational transition of SERCA, assessed through a Bcl-2-dependent increase in SERCA thiols available for the labelling with a fluorescent reagent. This partial unfolding of SERCA did not lead to a higher sensitivity of SERCA towards oxidative inactivation. Our results suggest that the direct interaction of Bcl-2 with SERCA may be involved in the regulation of apoptotic processes in vivo through modulation of cytoplasmic and/or endoplasmic reticulum calcium levels required for the execution of apoptosis.
Figures







Similar articles
-
Displacement of SERCA from SR lipid caveolae-related domains by Bcl-2: a possible mechanism for SERCA inactivation.Biochemistry. 2006 Jan 10;45(1):175-84. doi: 10.1021/bi050800s. Biochemistry. 2006. PMID: 16388593
-
Ca2+-dependent redox modulation of SERCA 2b by ERp57.J Cell Biol. 2004 Jan 5;164(1):35-46. doi: 10.1083/jcb.200307010. Epub 2003 Dec 29. J Cell Biol. 2004. PMID: 14699087 Free PMC article.
-
Heat-shock proteins attenuate SERCA inactivation by the anti-apoptotic protein Bcl-2: possible implications for the ER Ca2+-mediated apoptosis.Biochem J. 2012 May 15;444(1):127-39. doi: 10.1042/BJ20111114. Biochem J. 2012. PMID: 22360692 Free PMC article.
-
Sarco/endoplasmic reticulum Ca2+ (SERCA)-pumps: link to heart beats and calcium waves.Cell Calcium. 1999 Apr;25(4):277-90. doi: 10.1054/ceca.1999.0032. Cell Calcium. 1999. PMID: 10456225 Review.
-
The regulation of sarco(endo)plasmic reticulum calcium-ATPases (SERCA).Can J Physiol Pharmacol. 2015 Oct;93(10):843-54. doi: 10.1139/cjpp-2014-0463. Epub 2015 Jan 19. Can J Physiol Pharmacol. 2015. PMID: 25730320 Review.
Cited by
-
Alpha-helical destabilization of the Bcl-2-BH4-domain peptide abolishes its ability to inhibit the IP3 receptor.PLoS One. 2013 Aug 30;8(8):e73386. doi: 10.1371/journal.pone.0073386. eCollection 2013. PLoS One. 2013. PMID: 24137498 Free PMC article.
-
Ethyl 2-amino-6-(3,5-dimethoxyphenyl)-4-(2-ethoxy-2-oxoethyl)-4H-chromene-3-carboxylate (CXL017): a novel scaffold that resensitizes multidrug resistant leukemia cells to chemotherapy.ACS Chem Biol. 2013 Feb 15;8(2):327-35. doi: 10.1021/cb300460f. Epub 2012 Nov 5. ACS Chem Biol. 2013. PMID: 23102022 Free PMC article.
-
Novel role of HAX-1 in ischemic injury protection involvement of heat shock protein 90.Circ Res. 2013 Jan 4;112(1):79-89. doi: 10.1161/CIRCRESAHA.112.279935. Epub 2012 Sep 14. Circ Res. 2013. PMID: 22982986 Free PMC article.
-
Quantitative mapping of oxidation-sensitive cysteine residues in SERCA in vivo and in vitro by HPLC-electrospray-tandem MS: selective protein oxidation during biological aging.Biochem J. 2006 Mar 15;394(Pt 3):605-15. doi: 10.1042/BJ20051214. Biochem J. 2006. PMID: 16307534 Free PMC article.
-
Glucosamine protects neonatal cardiomyocytes from ischemia-reperfusion injury via increased protein O-GlcNAc and increased mitochondrial Bcl-2.Am J Physiol Cell Physiol. 2008 Jun;294(6):C1509-20. doi: 10.1152/ajpcell.00456.2007. Epub 2008 Mar 26. Am J Physiol Cell Physiol. 2008. PMID: 18367586 Free PMC article.
References
-
- Hajnoczky G., Csordas G., Yi M. Old players in a new role: mitochondria-associated membranes, VDAC, and ryanodine receptors as contributors to calcium signal propagation from endoplasmic reticulum to the mitochondria. Cell Calcium. 2002;32:363–377. - PubMed
-
- Green D. R., Reed J. C. Mitochondria and apoptosis. Science. 1998;281:1309–1312. - PubMed
-
- Martinou J. C., Green D. R. Breaking the mitochondrial barrier. Nat. Rev. Mol. Cell. Biol. 2001;2:63–67. - PubMed
-
- Berridge M. J., Lipp P., Bootman M. D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell. Biol. 2000;1:11–21. - PubMed
-
- Berridge M. J. Cell signalling. A tale of two messengers. Nature (London) 1993;361:315–325. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous