

Borealin: a novel chromosomal passenger required for stability of the bipolar mitotic spindle
- PMID: 15249581
- PMCID: PMC2172304
- DOI: 10.1083/jcb.200404001
Borealin: a novel chromosomal passenger required for stability of the bipolar mitotic spindle
Abstract
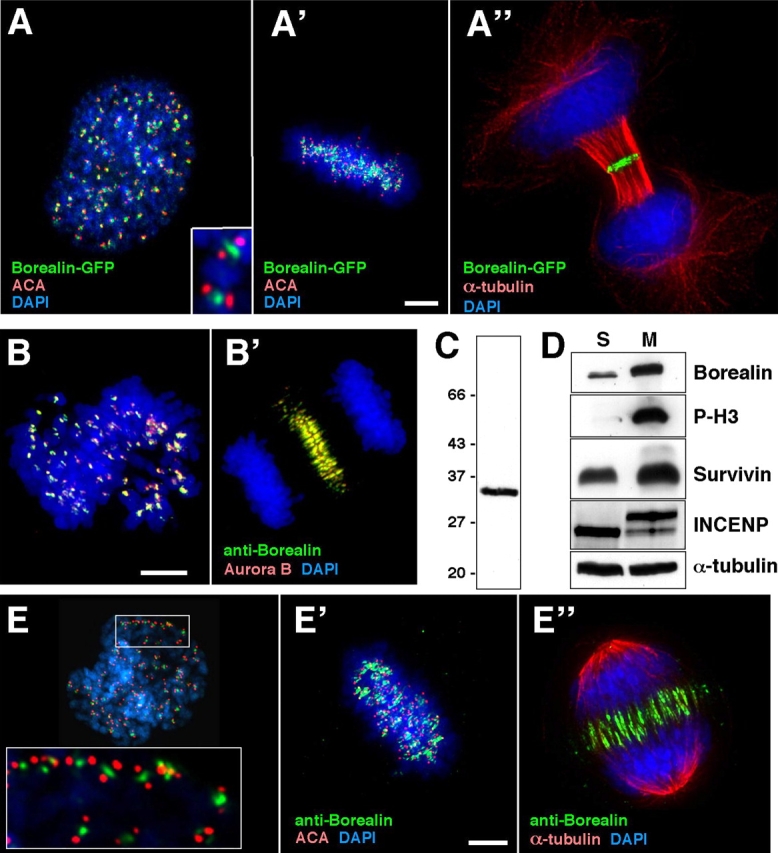
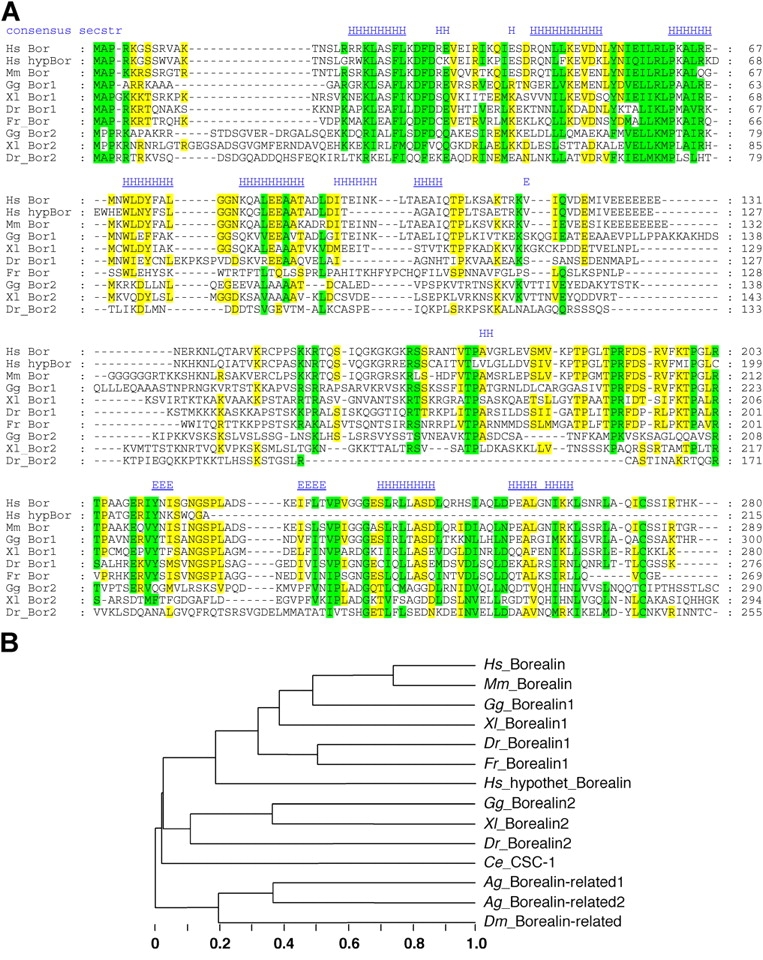
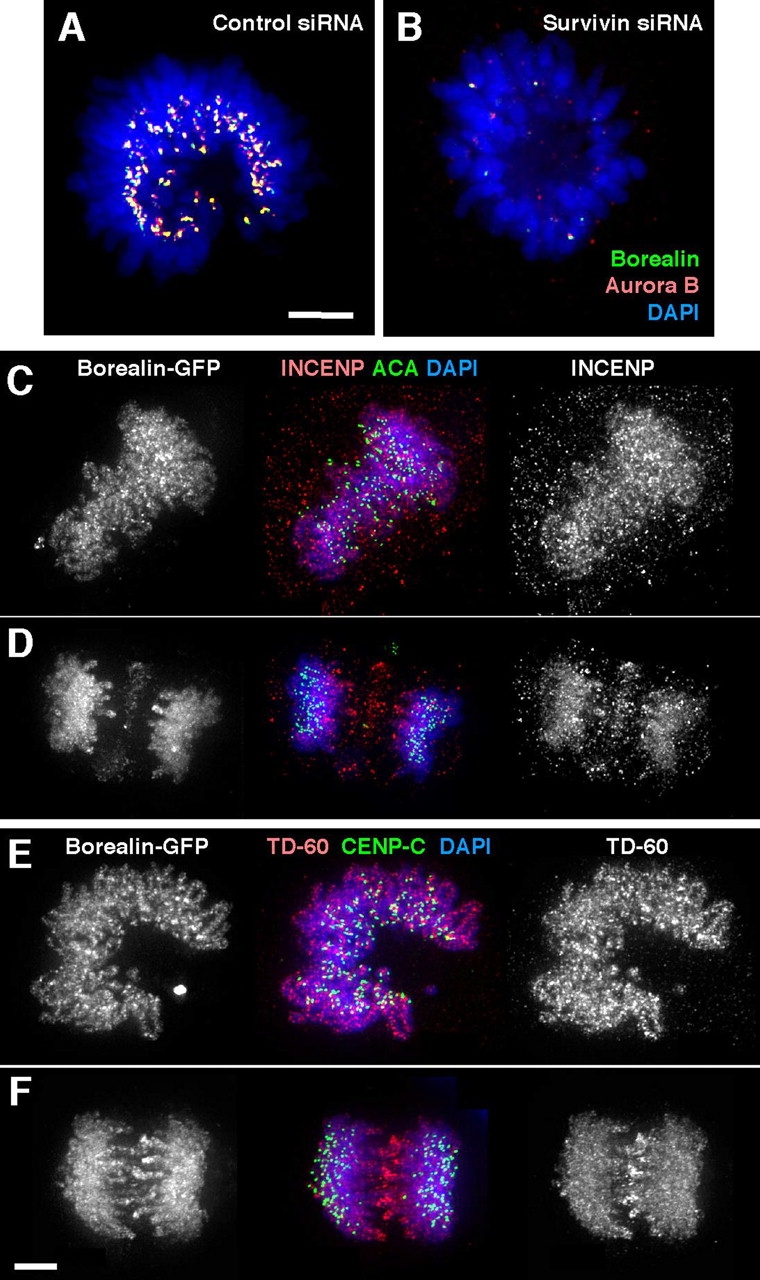
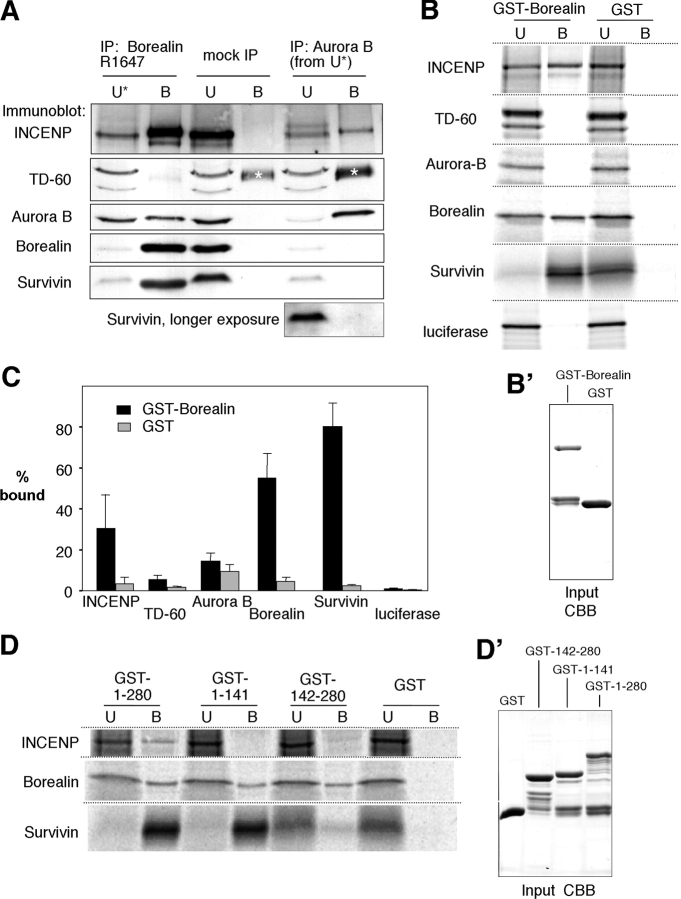
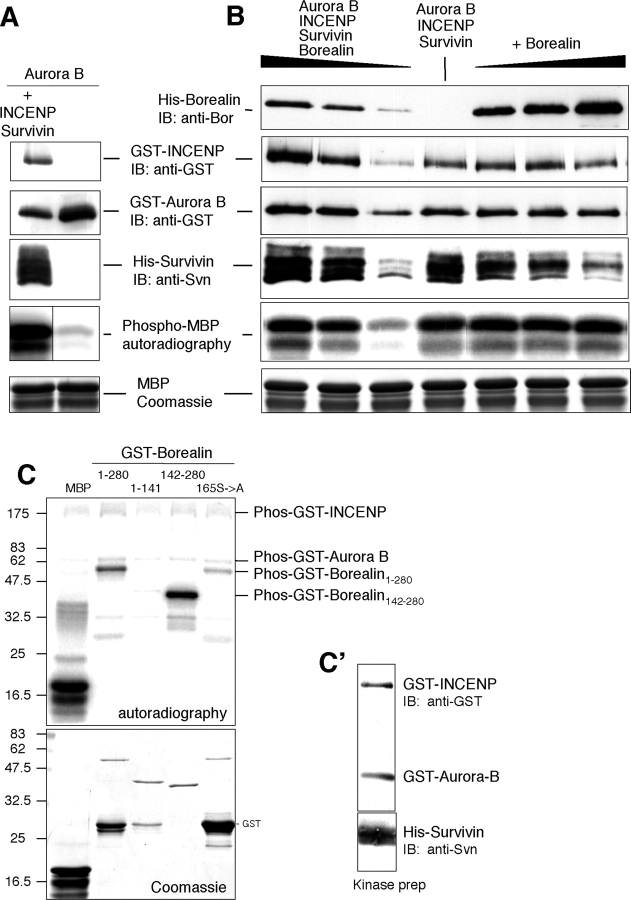
The chromosomal passenger complex of Aurora B kinase, INCENP, and Survivin has essential regulatory roles at centromeres and the central spindle in mitosis. Here, we describe Borealin, a novel member of the complex. Approximately half of Aurora B in mitotic cells is complexed with INCENP, Borealin, and Survivin; and Borealin binds Survivin and INCENP in vitro. A second complex contains Aurora B and INCENP, but no Borealin or Survivin. Depletion of Borealin by RNA interference delays mitotic progression and results in kinetochore-spindle misattachments and an increase in bipolar spindles associated with ectopic asters. The extra poles, which apparently form after chromosomes achieve a bipolar orientation, severely disrupt the partitioning of chromosomes in anaphase. Borealin depletion has little effect on histone H3 serine10 phosphorylation. These results implicate the chromosomal passenger holocomplex in the maintenance of spindle integrity and suggest that histone H3 serine10 phosphorylation is performed by an Aurora B-INCENP subcomplex.
Copyright The Rockerfeller University Press
Figures
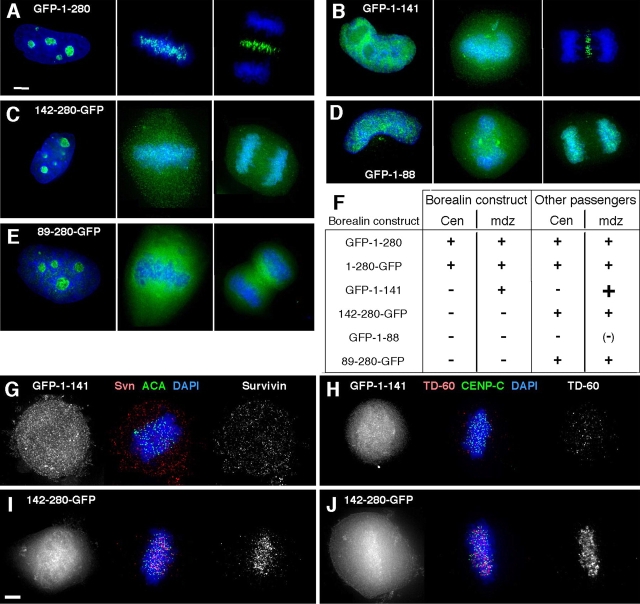
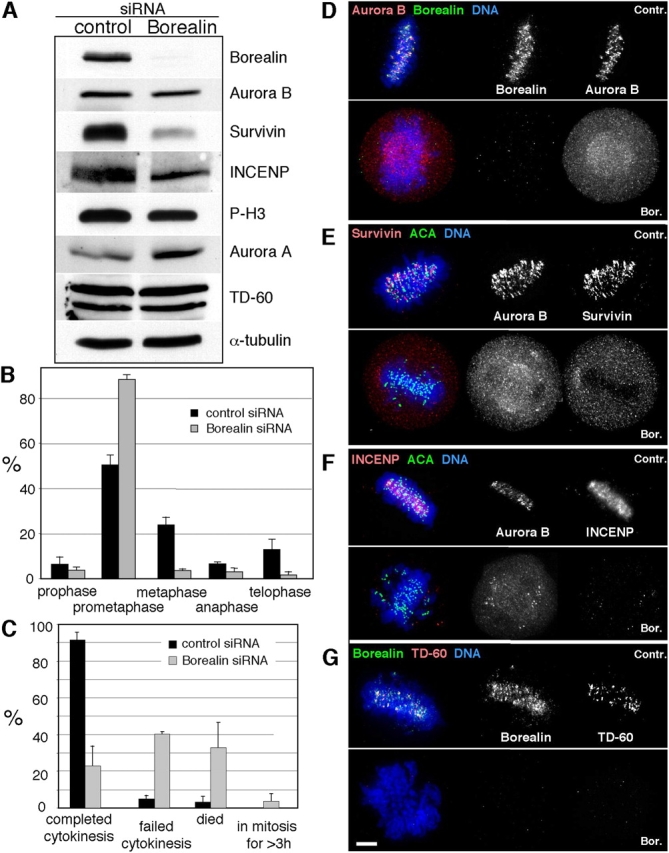
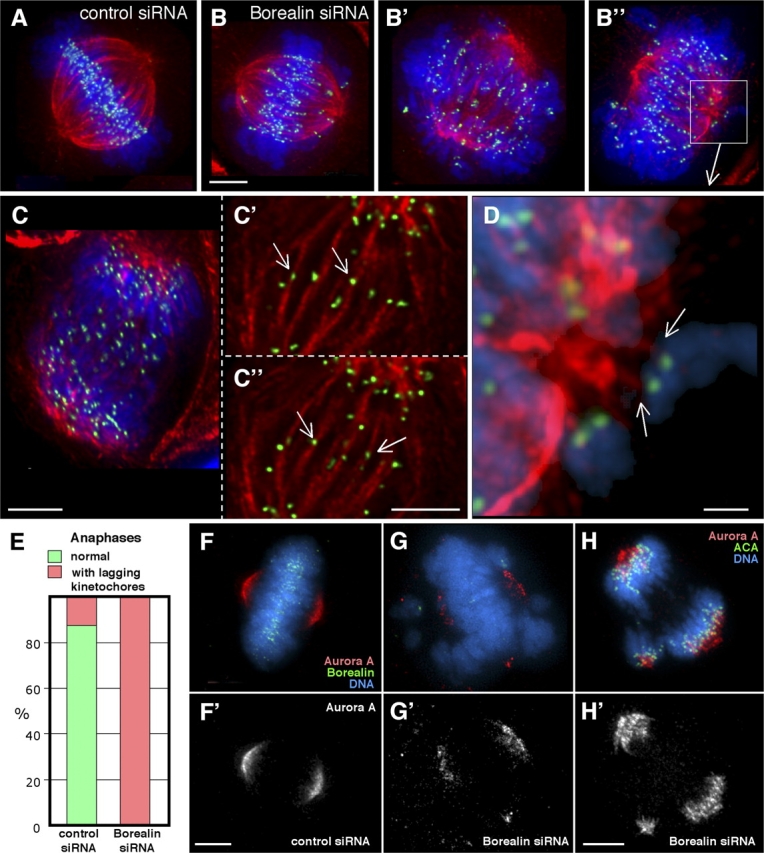
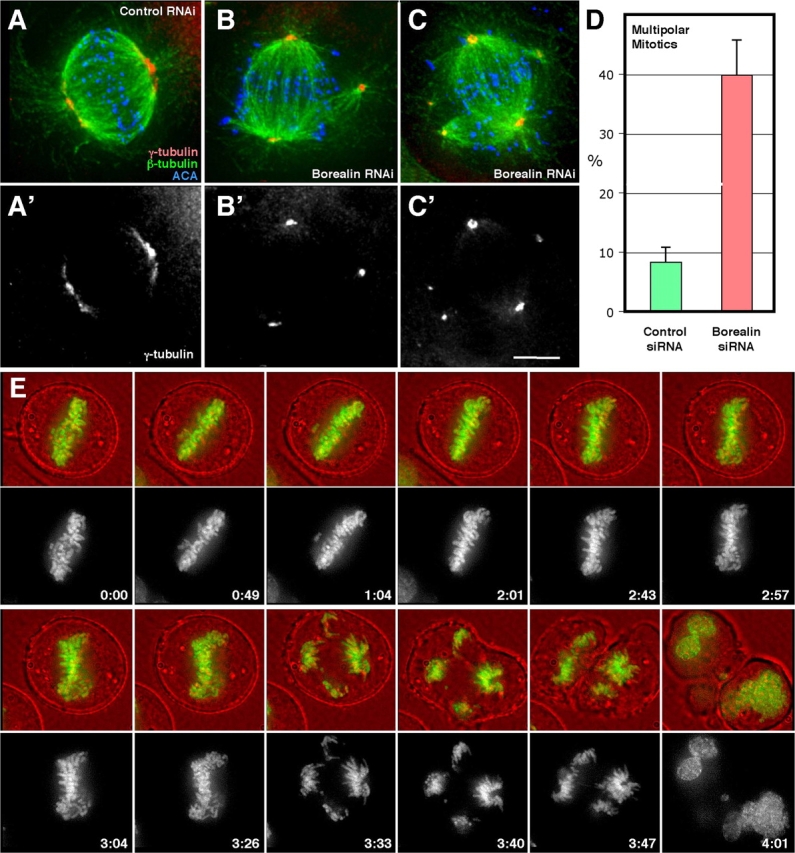
References
-
- Adams, R.R., S.P. Wheatley, A.M. Gouldsworthy, S.E. Kandels-Lewis, M. Carmena, C. Smythe, D.L. Gerloff, and W.C. Earnshaw. 2000. INCENP binds the aurora-related kinase AIRK2 and is required to target it to chromosomes, the central spindle and cleavage furrow. Curr. Biol. 10:1075–1078. - PubMed
-
- Andreassen, P.R., D.K. Palmer, M.H. Wener, and R.L. Margolis. 1991. Telophase disk: a new mammalian mitotic organelle that bisects telophase cells with a possible function in cytokinesis. J. Cell Sci. 99:523–534. - PubMed
-
- Andrews, P.D., E. Knatko, W.J. Moore, and J.R. Swedlow. 2003. Mitotic mechanics: the auroras come into view. Curr. Opin. Cell Biol. 15:672–683. - PubMed
-
- Andrews, P.D., Y. Ovechkina, N. Morrice, M. Wagenbach, K. Duncan, L. Wordeman, and J.R. Swedlow. 2004. Aurora B regulates MCAK at the mitotic centromere. Dev. Cell. 6:253–268. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
