

Functional implications of seizure-induced neurogenesis
- PMID: 15250595
- PMCID: PMC1839060
- DOI: 10.1007/978-1-4757-6376-8_14
Functional implications of seizure-induced neurogenesis
Abstract
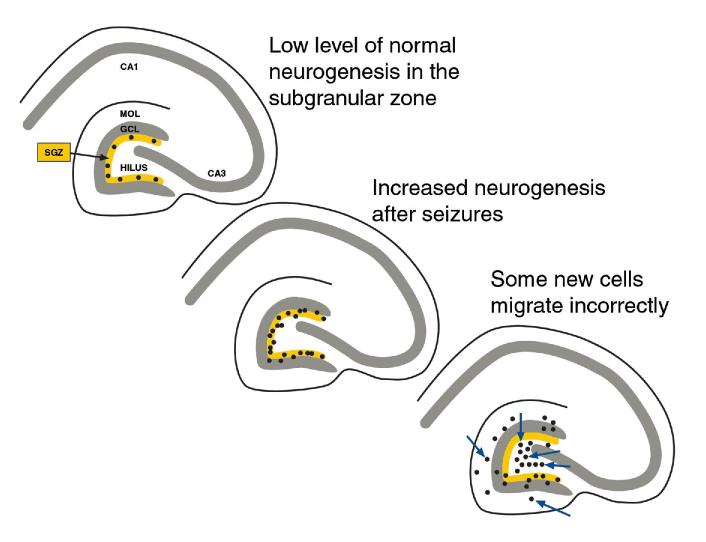
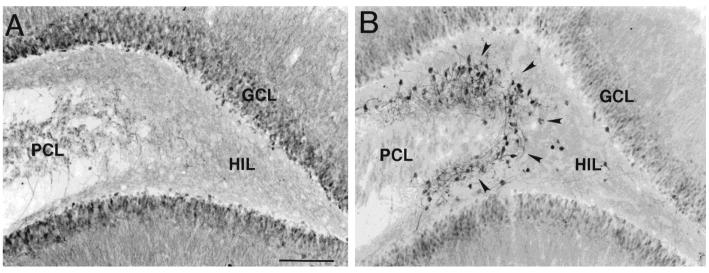
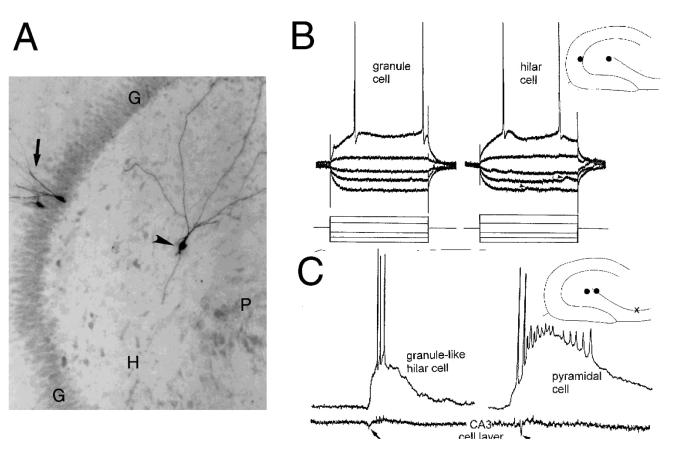
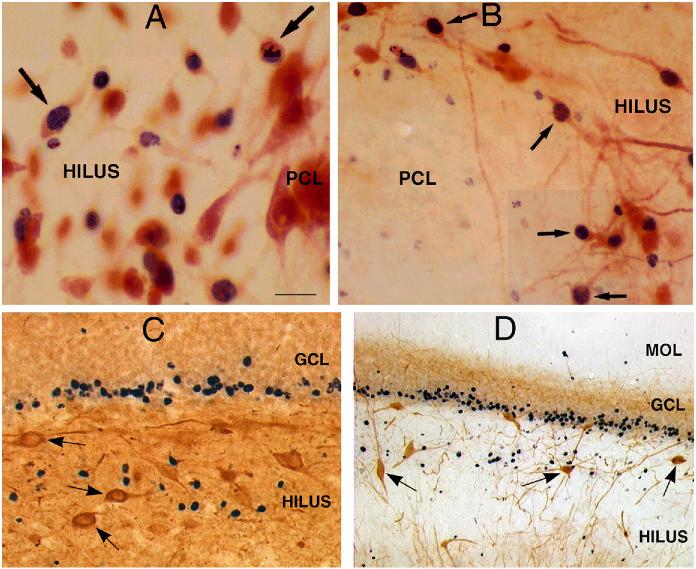
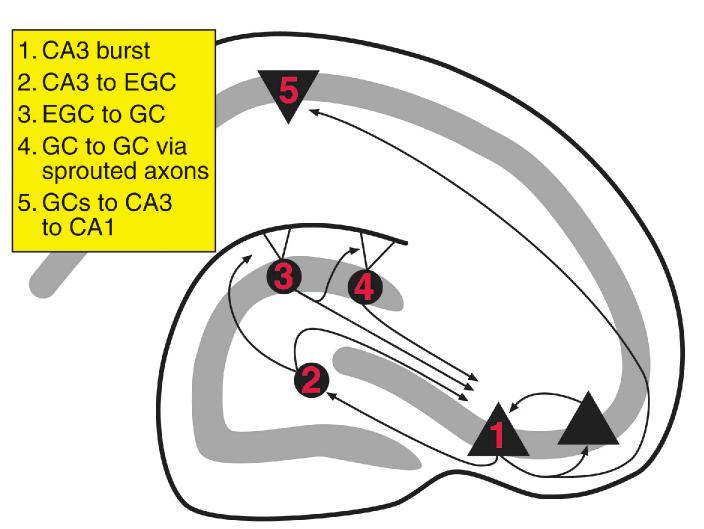
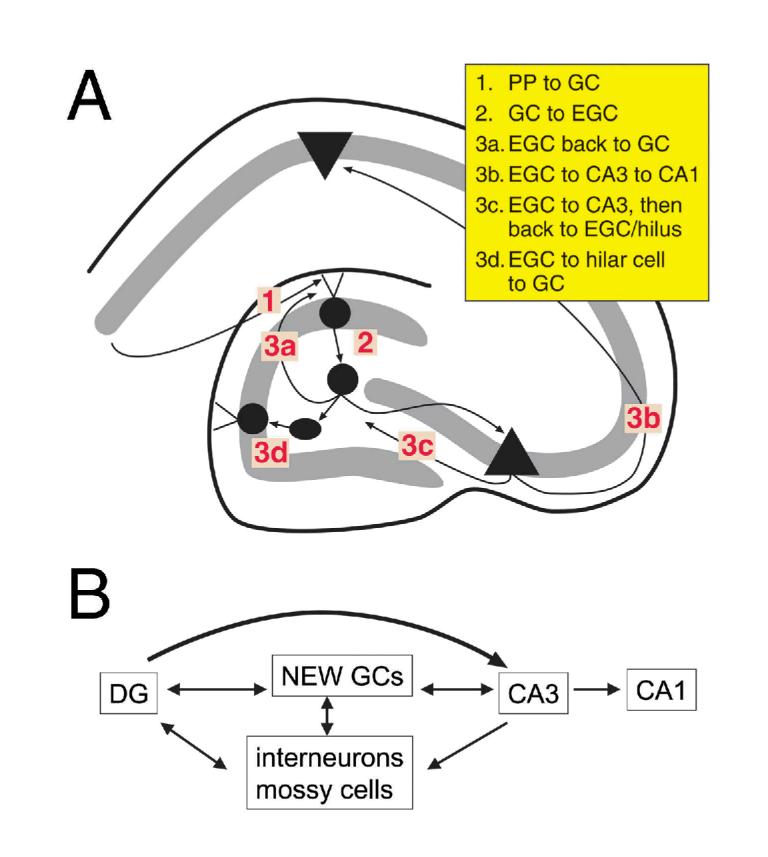
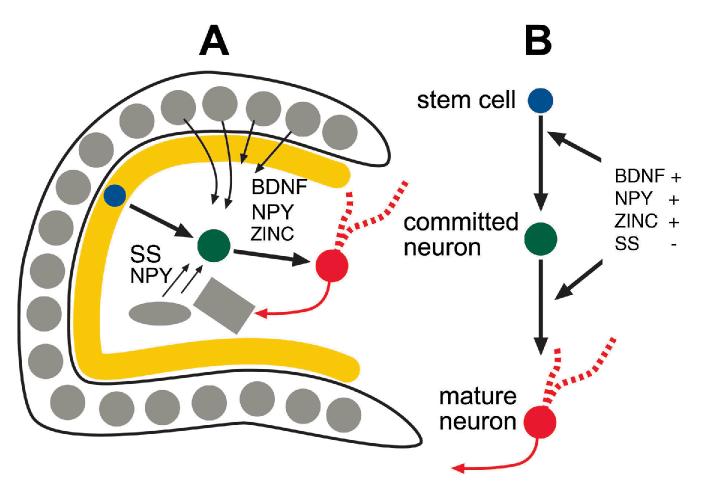
The neurobiological doctrine governing the concept of neurogenesis has undergone a revolution in the past few years. What was once considered dubious is now well accepted: new neurons are born in the adult brain. Science fiction is quickly becoming a reality as scientists discover ways to convert skin, bone, or blood cells into neurons. In the epilepsy arena, widespread interest has developed because of the evidence that neurogenesis increases after seizures, trauma, and other insults or injuries that alter seizure susceptibility. This review discusses some of the initial studies in this field, and their often surprising functional implications. The emphasis will be on the granule cells of hippocampus, because they are perhaps more relevant to epilepsy than other areas in which neurogenesis occurs throughout life, the olfactory bulb and subventricular zone. In particular, the following questions will be addressed: 1. Do granule cells that are born in the adult brain become functional, and what are the limits of their function? Do they behave homogeneously? Results from our own laboratory have focused on cells that become established outside the normal boundaries of the granule cell layer, forming a group of "ectopic" granule cells in the hilar region. 2. Is increased neurogenesis beneficial, or might it actually exacerbate seizures? Evidence is presented that supports the hypothesis that new granule cells may not necessarily act to ameliorate seizures, and might even contribute to them. Furthermore, cognitive deficits following seizures might in part be due to new circuits that develop between new cells and the host brain. 3. How do the new cells interact with the host brain? Several changes occur in the dentate gyrus after seizures, and increased neurogenesis is only one of many. What is the interdependence of this multitude of changes, if any? 4. Is neurogenesis increased after seizures in man? Research suggests that the data from human epileptics are actually inconsistent with the studies in animal models of epilepsy, because there is little evidence of increased neurogenesis in epileptic tissue resected from intractable epileptics. Yet neurogenesis has been shown to occur in humans throughout adult life. What might be the reasons for these seemingly disparate results?
Figures
Similar articles
-
Hippocampal transplants of fetal GABAergic progenitors regulate adult neurogenesis in mice with temporal lobe epilepsy.Neurobiol Dis. 2022 Nov;174:105879. doi: 10.1016/j.nbd.2022.105879. Epub 2022 Sep 29. Neurobiol Dis. 2022. PMID: 36183946 Free PMC article.
-
Seizure-induced neurogenesis: are more new neurons good for an adult brain?Prog Brain Res. 2002;135:121-31. doi: 10.1016/S0079-6123(02)35012-X. Prog Brain Res. 2002. PMID: 12143334 Review.
-
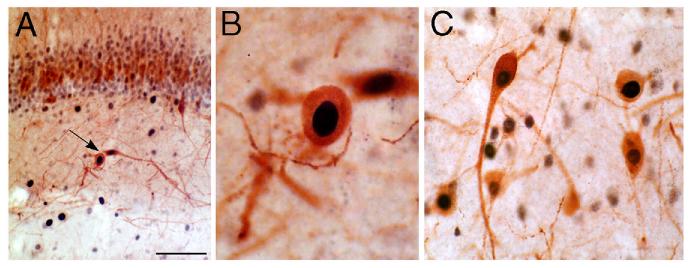
Granule-like neurons at the hilar/CA3 border after status epilepticus and their synchrony with area CA3 pyramidal cells: functional implications of seizure-induced neurogenesis.J Neurosci. 2000 Aug 15;20(16):6144-58. doi: 10.1523/JNEUROSCI.20-16-06144.2000. J Neurosci. 2000. PMID: 10934264 Free PMC article.
-
Adult neurogenesis in the intact and epileptic dentate gyrus.Prog Brain Res. 2007;163:529-40. doi: 10.1016/S0079-6123(07)63028-3. Prog Brain Res. 2007. PMID: 17765736 Review.
-
Neuroplasticity in the damaged dentate gyrus of the epileptic brain.Prog Brain Res. 2002;136:319-28. doi: 10.1016/s0079-6123(02)36027-8. Prog Brain Res. 2002. PMID: 12143392 Review.
Cited by
-
Adult neurogenesis in the mouse dentate gyrus protects the hippocampus from neuronal injury following severe seizures.Hippocampus. 2019 Aug;29(8):683-709. doi: 10.1002/hipo.23062. Epub 2019 Jan 23. Hippocampus. 2019. PMID: 30672046 Free PMC article.
-
The Dentate Gyrus and Temporal Lobe Epilepsy: An "Exciting" Era.Epilepsy Curr. 2019 Jul-Aug;19(4):249-255. doi: 10.1177/1535759719855952. Epub 2019 Jun 24. Epilepsy Curr. 2019. PMID: 31232111 Free PMC article.
-
Hippocampal transplants of fetal GABAergic progenitors regulate adult neurogenesis in mice with temporal lobe epilepsy.Neurobiol Dis. 2022 Nov;174:105879. doi: 10.1016/j.nbd.2022.105879. Epub 2022 Sep 29. Neurobiol Dis. 2022. PMID: 36183946 Free PMC article.
-
Novel Psychopharmacological Herbs Relieve Behavioral Abnormalities and Hippocampal Dysfunctions in an Animal Model of Post-Traumatic Stress Disorder.Nutrients. 2023 Aug 31;15(17):3815. doi: 10.3390/nu15173815. Nutrients. 2023. PMID: 37686847 Free PMC article.
-
Epilepsy as a Network Disorder (2): What can we learn from other network disorders such as dementia and schizophrenia, and what are the implications for translational research?Epilepsy Behav. 2018 Jan;78:302-312. doi: 10.1016/j.yebeh.2017.09.016. Epub 2017 Oct 31. Epilepsy Behav. 2018. PMID: 29097123 Free PMC article. Review.
References
-
- Altman J, Bayer SA. Migration and distribution of two populations of hippocampal granule cell precursors during perinatal and postnatal periods. J Comp Neurol. 1990;301:365–381. - PubMed
-
- Altman J, Bayer SA. Mosaic organization of the hippocampal neuroepithelium and the multiple germinal sources of dentate granule cells. J Comp Neurol. 1990;301:325–342. - PubMed
-
- Amaral DG. A Golgi study of cell types in the hilar region of the hippocampus in the rat. J Comp Neurol. 1978;182:851–914. - PubMed
-
- Azizi SA. Exploiting nonneuronal cells to rebuild the nervous system: from bone marrow to brain. The Neuroscientist. 2000;6:353–361.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
