

La autoantigen is necessary for optimal function of the poliovirus and hepatitis C virus internal ribosome entry site in vivo and in vitro
- PMID: 15254251
- PMCID: PMC444852
- DOI: 10.1128/MCB.24.15.6861-6870.2004
La autoantigen is necessary for optimal function of the poliovirus and hepatitis C virus internal ribosome entry site in vivo and in vitro
Abstract
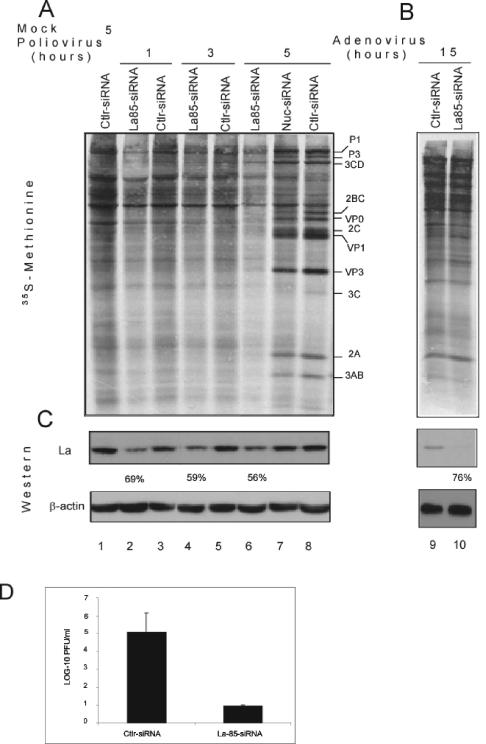
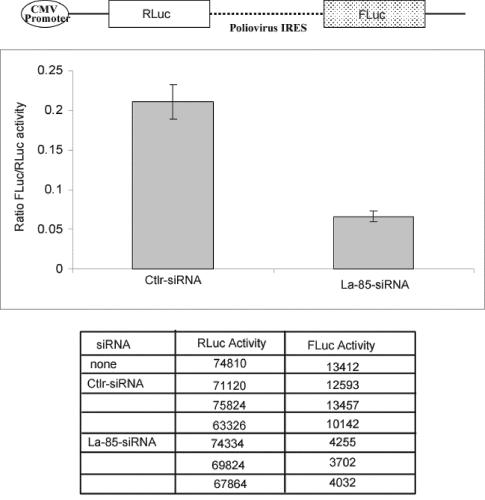
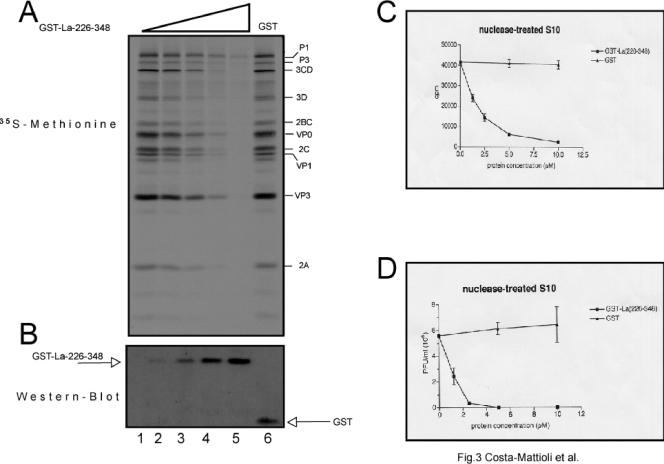
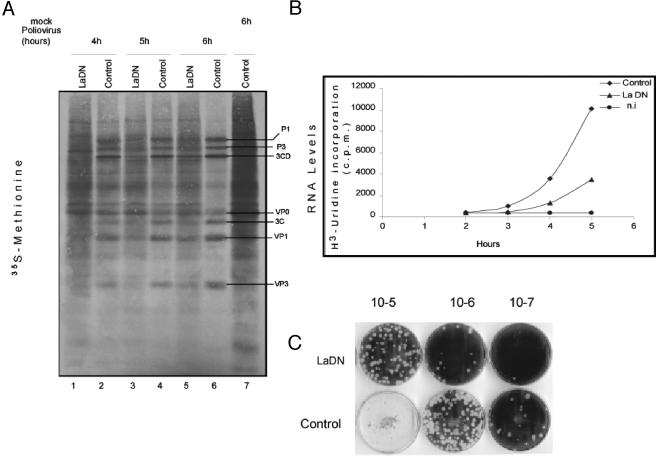
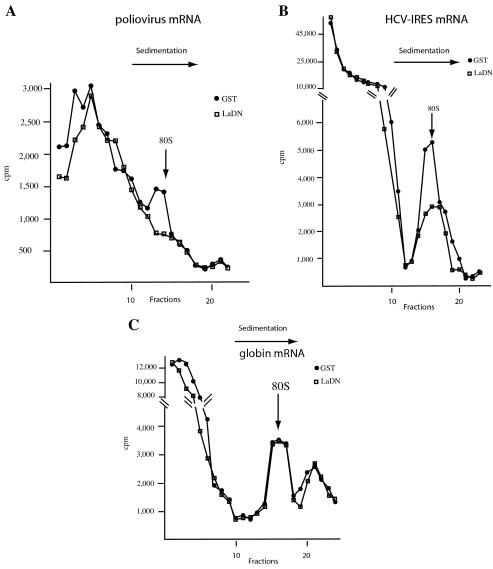
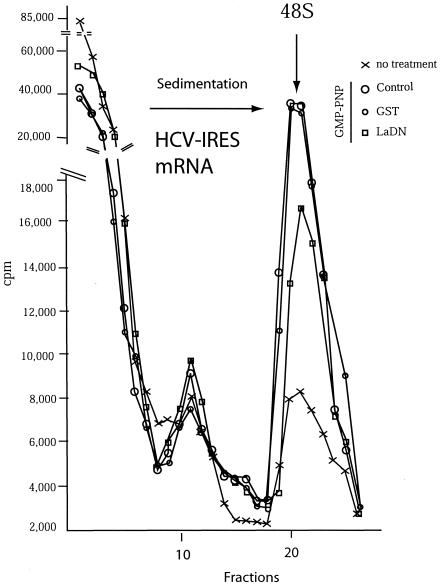
Translation of poliovirus and hepatitis C virus (HCV) RNAs is initiated by recruitment of 40S ribosomes to an internal ribosome entry site (IRES) in the mRNA 5' untranslated region. Translation initiation of these RNAs is stimulated by noncanonical initiation factors called IRES trans-activating factors (ITAFs). The La autoantigen is such an ITAF, but functional evidence for the role of La in poliovirus and HCV translation in vivo is lacking. Here, by two methods using small interfering RNA and a dominant-negative mutant of La, we demonstrate that depletion of La causes a dramatic reduction in poliovirus IRES function in vivo. We also show that 40S ribosomal subunit binding to HCV and poliovirus IRESs in vitro is inhibited by a dominant-negative form of La. These results provide strong evidence for a function of the La autoantigen in IRES-dependent translation and define the step of translation which is stimulated by La.
Figures
Similar articles
-
Lower concentration of La protein required for internal ribosome entry on hepatitis C virus RNA than on poliovirus RNA.J Gen Virol. 1999 Sep;80 ( Pt 9):2319-2327. doi: 10.1099/0022-1317-80-9-2319. J Gen Virol. 1999. PMID: 10501483
-
Targeting internal ribosome entry site (IRES)-mediated translation to block hepatitis C and other RNA viruses.FEMS Microbiol Lett. 2004 May 15;234(2):189-99. doi: 10.1016/j.femsle.2004.03.045. FEMS Microbiol Lett. 2004. PMID: 15135522 Review.
-
A peptide derived from RNA recognition motif 2 of human la protein binds to hepatitis C virus internal ribosome entry site, prevents ribosomal assembly, and inhibits internal initiation of translation.J Virol. 2005 Aug;79(15):9842-53. doi: 10.1128/JVI.79.15.9842-9853.2005. J Virol. 2005. PMID: 16014945 Free PMC article.
-
Structural determinant of human La protein critical for internal initiation of translation of hepatitis C virus RNA.J Virol. 2008 Dec;82(23):11927-38. doi: 10.1128/JVI.00924-08. Epub 2008 Oct 1. J Virol. 2008. PMID: 18829760 Free PMC article.
-
Structural and mechanistic insights into hepatitis C viral translation initiation.Nat Rev Microbiol. 2007 Jan;5(1):29-38. doi: 10.1038/nrmicro1558. Epub 2006 Nov 27. Nat Rev Microbiol. 2007. PMID: 17128284 Review.
Cited by
-
Nuclear proteins hijacked by mammalian cytoplasmic plus strand RNA viruses.Virology. 2015 May;479-480:457-74. doi: 10.1016/j.virol.2015.03.001. Epub 2015 Mar 26. Virology. 2015. PMID: 25818028 Free PMC article. Review.
-
Giardiavirus internal ribosome entry site has an apparently unique mechanism of initiating translation.PLoS One. 2009 Oct 14;4(10):e7435. doi: 10.1371/journal.pone.0007435. PLoS One. 2009. PMID: 19826476 Free PMC article.
-
N-Terminomics TAILS Identifies Host Cell Substrates of Poliovirus and Coxsackievirus B3 3C Proteinases That Modulate Virus Infection.J Virol. 2018 Mar 28;92(8):e02211-17. doi: 10.1128/JVI.02211-17. Print 2018 Apr 15. J Virol. 2018. PMID: 29437971 Free PMC article.
-
Human Cytomegalovirus Strategies to Maintain and Promote mRNA Translation.Viruses. 2016 Apr 13;8(4):97. doi: 10.3390/v8040097. Viruses. 2016. PMID: 27089357 Free PMC article. Review.
-
Evidence that PTB does not stimulate HCV IRES-driven translation.Virus Genes. 2007 Aug;35(1):5-15. doi: 10.1007/s11262-006-0038-z. Epub 2006 Oct 13. Virus Genes. 2007. PMID: 17039407
References
-
- Ali, N., G. J. Pruijn, D. J. Kenan, J. D. Keene, and A. Siddiqui. 2000. Human La antigen is required for the hepatitis C virus internal ribosome entry site-mediated translation. J. Biol. Chem. 275:27531-27540. - PubMed
-
- Bartenschlager, R. 2002. Hepatitis C virus replicons: potential role for drug development. Nat. Rev. Drug Discov. 1:911-916. - PubMed
-
- Belsham, G. J., and R. J. Jackson. 2000. Translation initiation on picornavirus RNA, p. 869-900. In N. Sonenberg, J. W. B. Hershey, and W. C. Merrick (ed.), Translational control of gene expression. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases