

doi: 10.1128/JB.186.15.4838-4843.2004.
Adaptive point mutation and adaptive amplification pathways in the Escherichia coli Lac system: stress responses producing genetic change
Affiliations
- PMID: 15262914
- PMCID: PMC451650
- DOI: 10.1128/JB.186.15.4838-4843.2004
Item in Clipboard
Adaptive point mutation and adaptive amplification pathways in the Escherichia coli Lac system: stress responses producing genetic change
J Bacteriol.
2004 Aug.
No abstract available
Figures
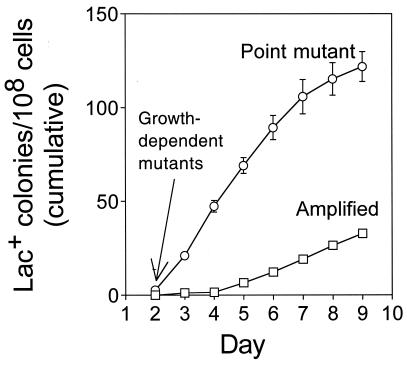
Adaptive point mutation and adaptive gene amplification are separate outcomes that arise with different kinetics. Point mutants have compensatory frameshift reversions in the lac gene (19, 61) and also carry high frequencies of other mutations genome wide (24, 57, 70). lac-amplified clones carry 20 to 50 tandem repeats of 7- to 40-kb DNA segments spanning the leaky lac frameshift allele, which allow growth on lactose medium without a compensatory frameshift mutation (30). lac-amplified clones do not carry high levels of extra mutations genome wide (30). Shown is a cumulative plot. The lac-amplified clones comprise typically ≥40% of new colonies from day 8 onward (30). Data are from reference .
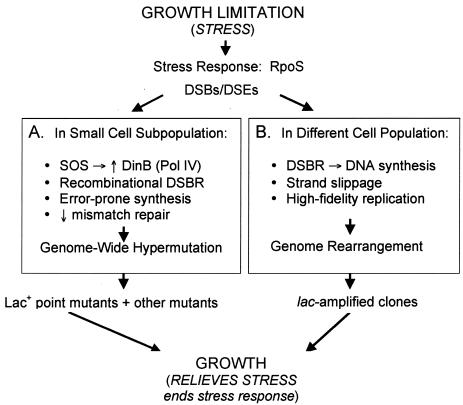
A stress-response (HM) model for adaptive point mutation and amplification (modified from references , , and 58). Starvation is proposed to promote RpoS-dependent stress responses that make the normally high-fidelity process of DNA double-strand-break repair (DSBR) error prone in two different ways. (A) Version I: in a small subpopulation of cells, genome-wide hypermutation is caused by high-level expression of the error-prone DNA polymerase DinB (Pol IV) (66, 71), which is upregulated both by the SOS response (37, 38) and also by RpoS (41), the stationary-phase and general stress-response transcription (σ) factor (35) required both for adaptive point mutation and amplification (45). Also in this population, mismatch repair becomes limiting transiently, perhaps via saturation by excess DNA polymerase errors (27) made by DinB (14, 49). The outcome is genome-wide hypermutation wherever DNA double-strand breaks (DSBs) or ends (DSEs) are formed and repaired. (Because more DSEs form in the F′ plasmid than in the chromosome, due to single-strand nicks made at the F transfer origin, there is more mutation on F′ [17] than on the chromosome [5], but the chromosome still experiences DSBR-protein- and DinB-dependent stationary-phase point mutations [5] because it still sometimes has DSEs to repair.) Version II: an alternative version (not drawn) of this idea, compatible with the hypothesis that two cell populations generate point mutations (57) (discussed in the text), is that both populations have a mutagenic stress response leading to increased DinB. (This is necessary because most point mutations [85%] are DinB dependent). However, one population is less mutable than the other only because it has less DinB, because, e.g., it experiences only RpoS, but not SOS, induction of DinB, whereas the other experiences both and makes more DinB (and then also saturates mismatch repair), making it more mutable. This alternative is more complicated than version I, but possible. (B) We suggest that amplification is provoked by error-prone DSBR of a different sort: that DNA synthesis primed during DSBR lacks the controls of replication from an origin and in starving cells is slower and more likely to stall due to limiting nucleotides. We suggest that upon stalling, template switching occurs (similar to recombination models discussed in reference ; see also reference 72). This can produce a novel junction sequence in the DNA that leads to amplification either via promoting rolling circle replication or generating a duplication that amplifies to many copies by recombination (illustrated in reference 31). Because adaptive amplification is SOS and DinB independent (49), the role played by RpoS in adaptive amplification cannot be to up-regulate DinB (45). RpoS may play different roles in point mutation and amplification (45).
Comment in
-
Rebuttal: adaptive point mutation (Rosenberg and Hastings).J Bacteriol. 2004 Aug;186(15):4844. doi: 10.1128/JB.186.15.4844.2004. J Bacteriol. 2004. PMID: 15262915 Free PMC article. No abstract available.
-
Rebuttal: adaptive point mutation (Rosenberg and Hastings).J Bacteriol. 2004 Aug;186(15):4845. doi: 10.1128/JB.186.15.4845.2004. J Bacteriol. 2004. PMID: 15262916 Free PMC article. No abstract available.
References
-
- Andersson, D. I., E. S. Slechta, and J. R. Roth. 1998. Evidence that gene amplification underlies adaptive mutability of the bacterial lac operon. Science 282:1133-1135. - PubMed
-
- Bjedov, I., O. Tenaillon, B. Gérard, V. Souza, E. Denamur, M. Radman, F. Taddei, and I. Matic. 2003. Stress-induced mutagenesis in bacteria. Science 300:1404-1409. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
