

A new human somatic stem cell from placental cord blood with intrinsic pluripotent differentiation potential
- PMID: 15263023
- PMCID: PMC2212008
- DOI: 10.1084/jem.20040440
A new human somatic stem cell from placental cord blood with intrinsic pluripotent differentiation potential
Abstract
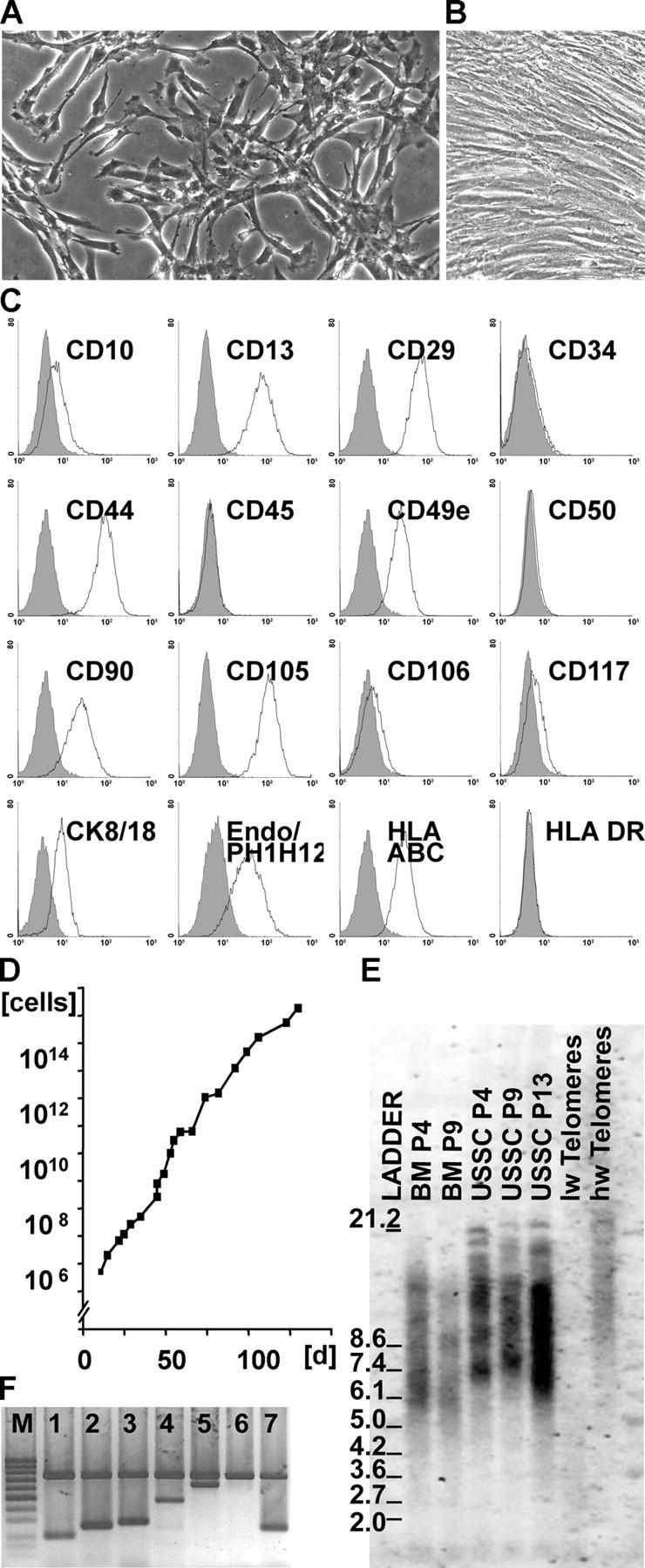
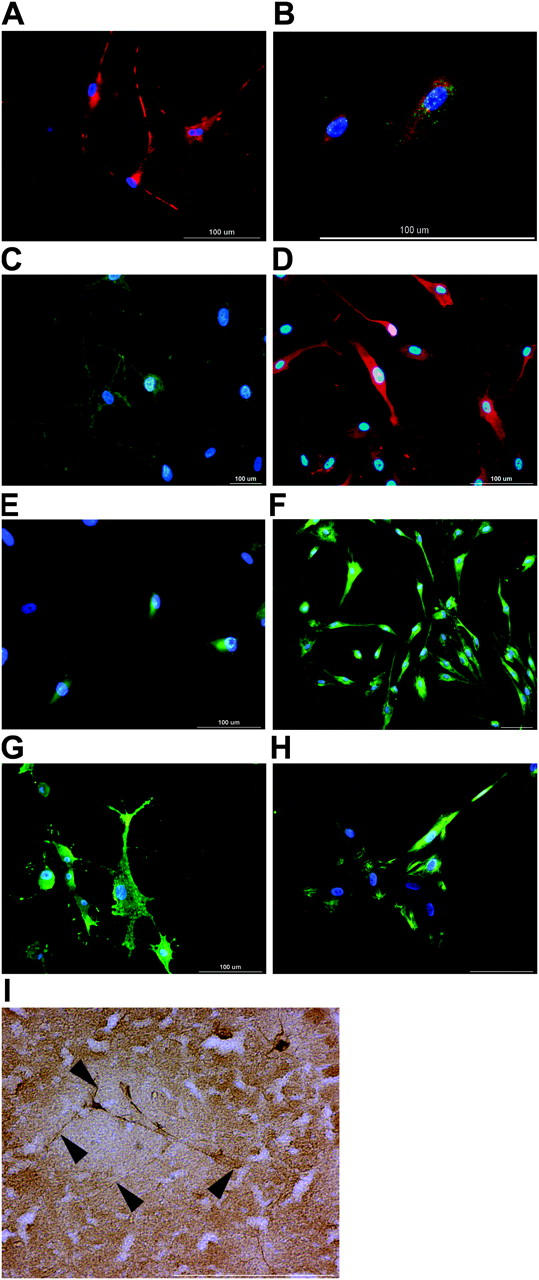
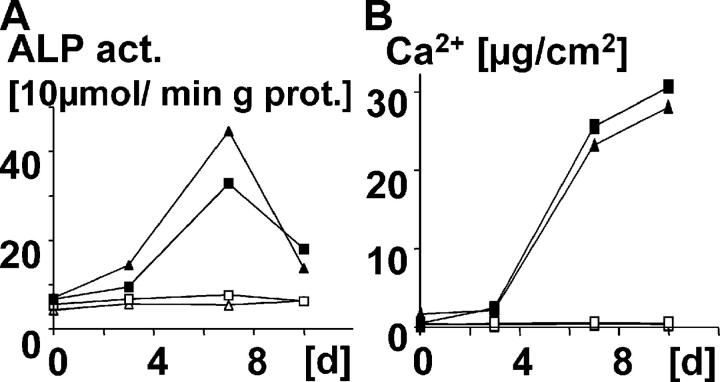
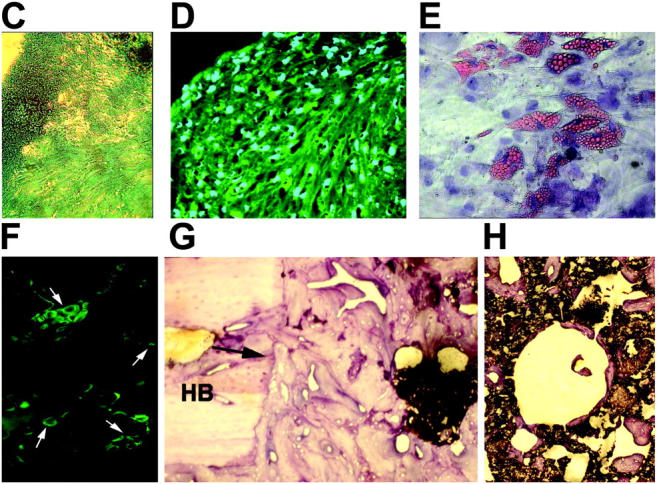
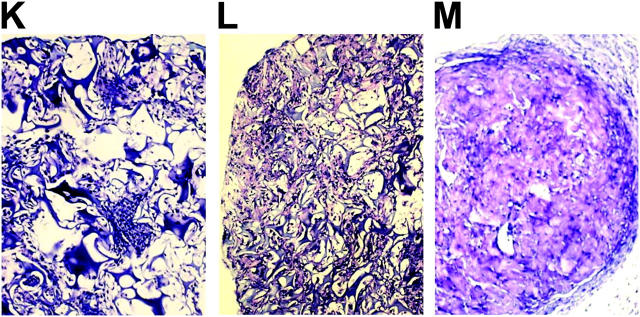
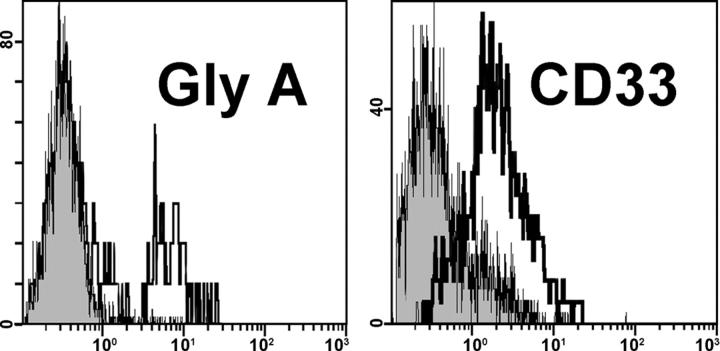
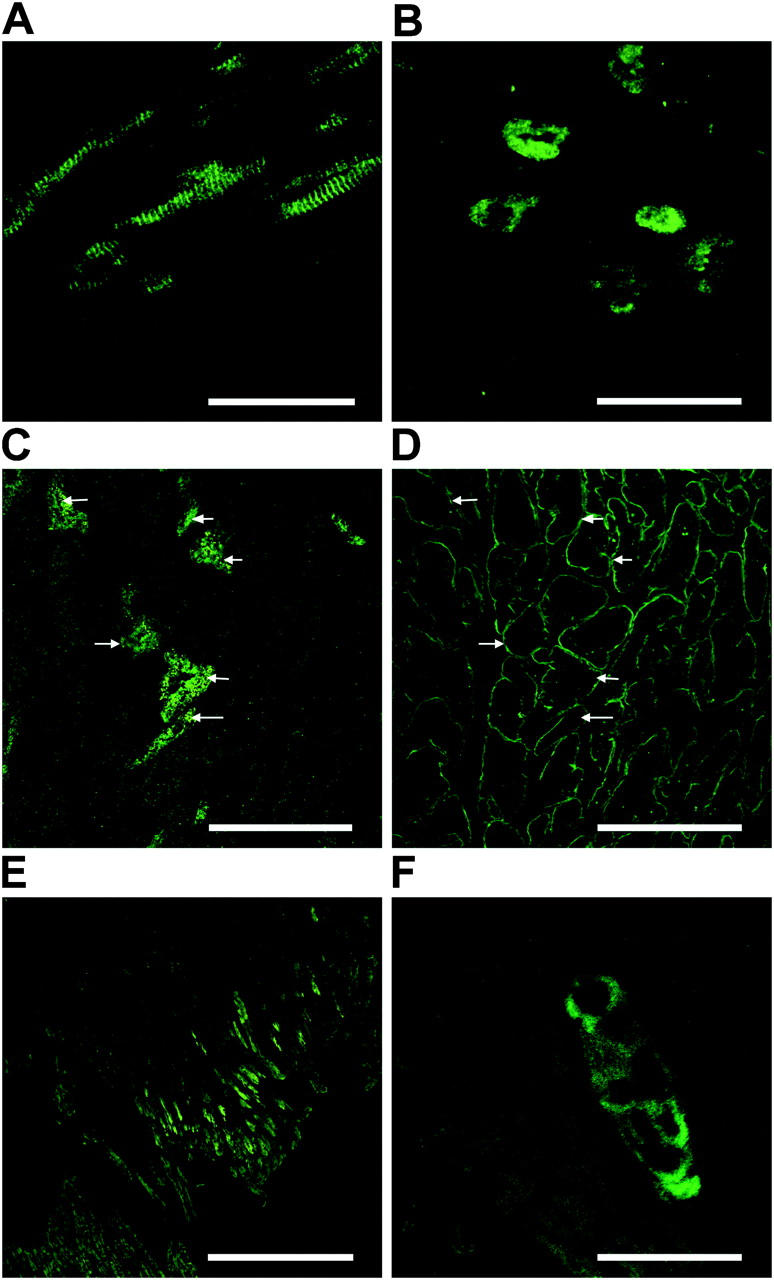
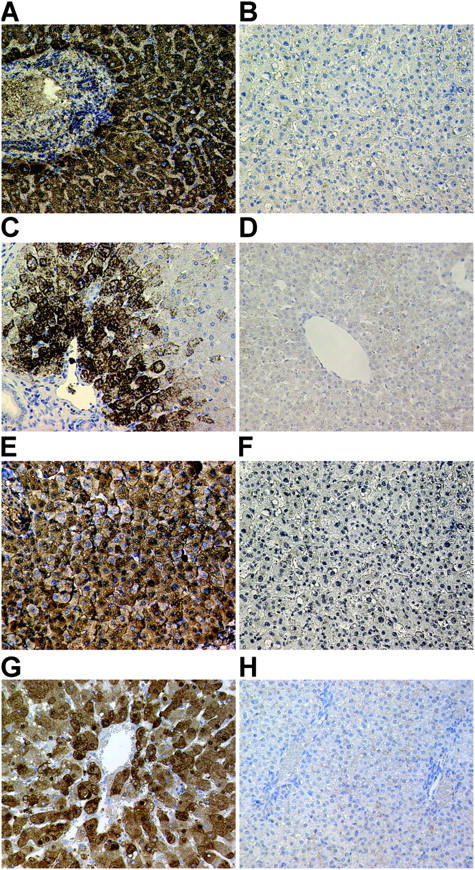
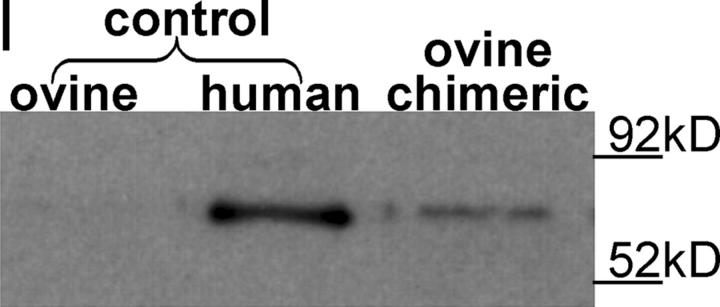
Here a new, intrinsically pluripotent, CD45-negative population from human cord blood, termed unrestricted somatic stem cells (USSCs) is described. This rare population grows adherently and can be expanded to 10(15) cells without losing pluripotency. In vitro USSCs showed homogeneous differentiation into osteoblasts, chondroblasts, adipocytes, and hematopoietic and neural cells including astrocytes and neurons that express neurofilament, sodium channel protein, and various neurotransmitter phenotypes. Stereotactic implantation of USSCs into intact adult rat brain revealed that human Tau-positive cells persisted for up to 3 mo and showed migratory activity and a typical neuron-like morphology. In vivo differentiation of USSCs along mesodermal and endodermal pathways was demonstrated in animal models. Bony reconstitution was observed after transplantation of USSC-loaded calcium phosphate cylinders in nude rat femurs. Chondrogenesis occurred after transplanting cell-loaded gelfoam sponges into nude mice. Transplantation of USSCs in a noninjury model, the preimmune fetal sheep, resulted in up to 5% human hematopoietic engraftment. More than 20% albumin-producing human parenchymal hepatic cells with absence of cell fusion and substantial numbers of human cardiomyocytes in both atria and ventricles of the sheep heart were detected many months after USSC transplantation. No tumor formation was observed in any of these animals.
Figures

References
-
- Thomson, J.A., J. Itskovitz-Eldor, S.S. Shapiro, M.A. Waknitz, J.J. Swiergiel, V.S. Marshall, and J.M. Jones. 1998. Embryonic stem cell lines derived from human blastocysts. Science. 282:1145–1147. - PubMed
-
- Erdo, F., C. Buhrle, J. Blunk, M. Hoehn, Y. Xia, B. Fleischmann, M. Focking, E. Kustermann, E. Kolossov, J. Hescheler, et al. 2003. Host-dependent tumorigenesis of embryonic stem cell transplantation in experimental stroke. J. Cereb. Blood Flow Metab. 23:780–785. - PubMed
-
- Gussoni, E., Y. Soneoka, C.D. Strickland, E.A. Buzney, M.K. Khan, A.F. Flint, L.M. Kunkel, and R.C. Mulligan. 1999. Dystrophin expression in the mdx mouse restored by stem cell transplantation. Nature. 401:390–394. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
