

A role for thymic stromal lymphopoietin in CD4(+) T cell development
- PMID: 15263024
- PMCID: PMC2212020
- DOI: 10.1084/jem.20031975
A role for thymic stromal lymphopoietin in CD4(+) T cell development
Abstract
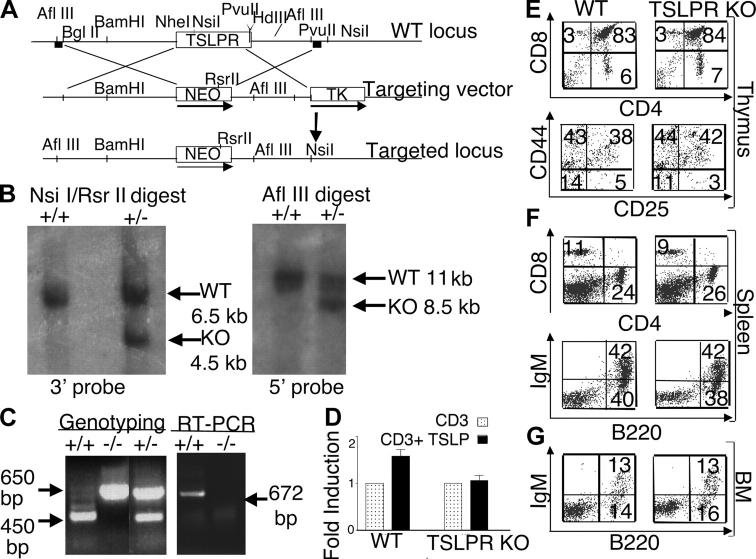
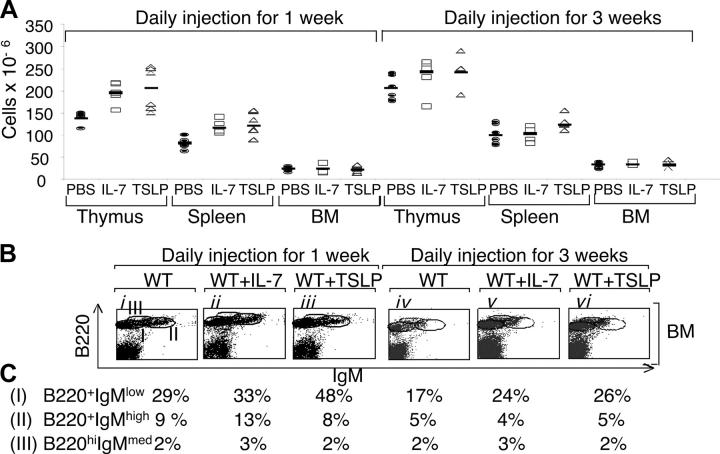
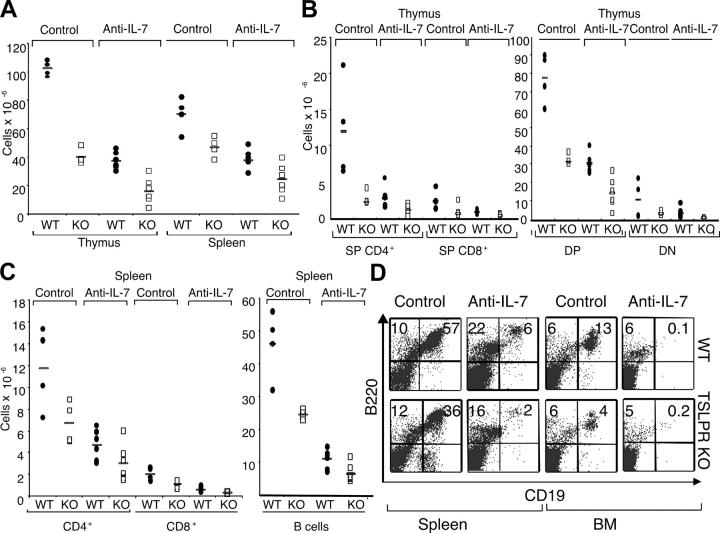
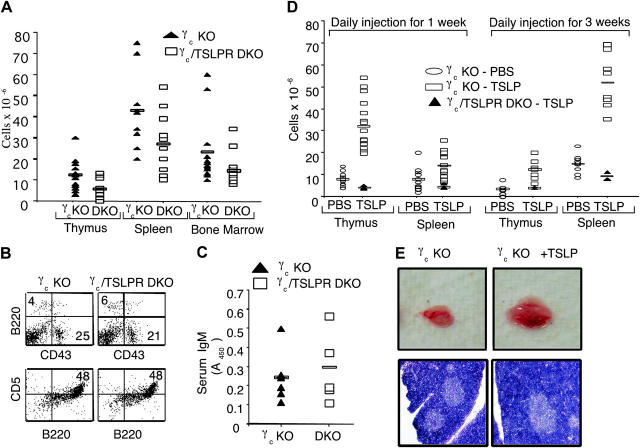
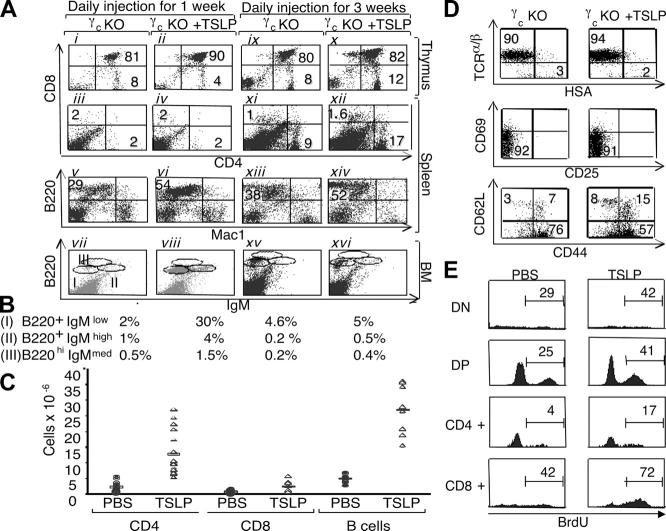
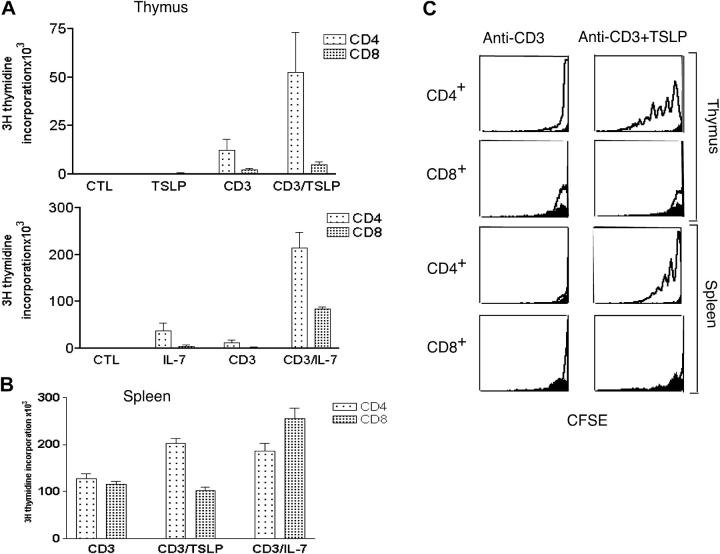
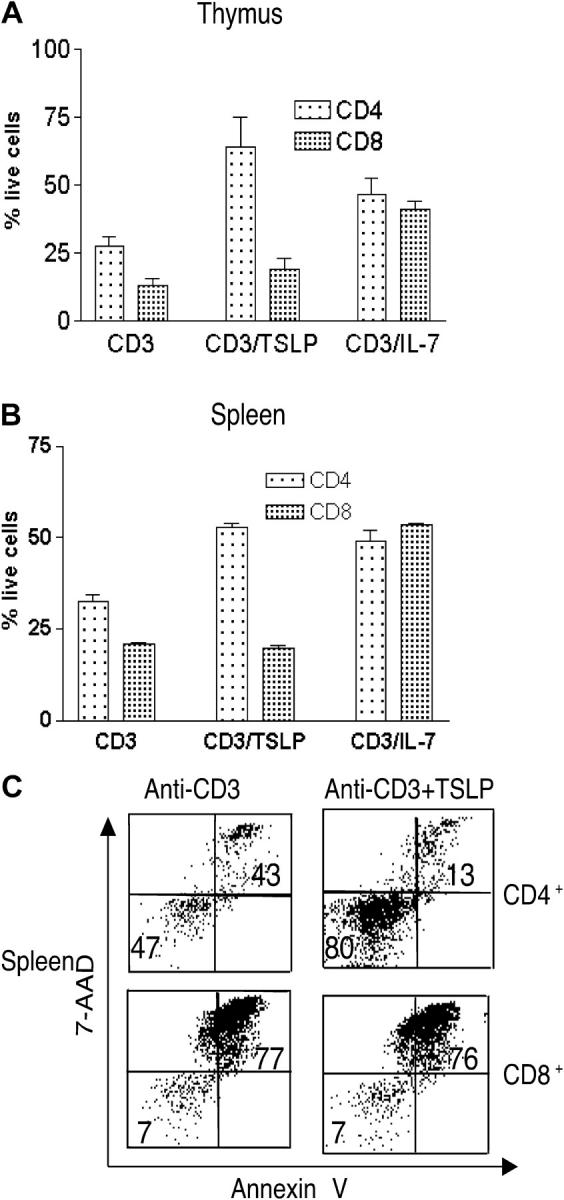
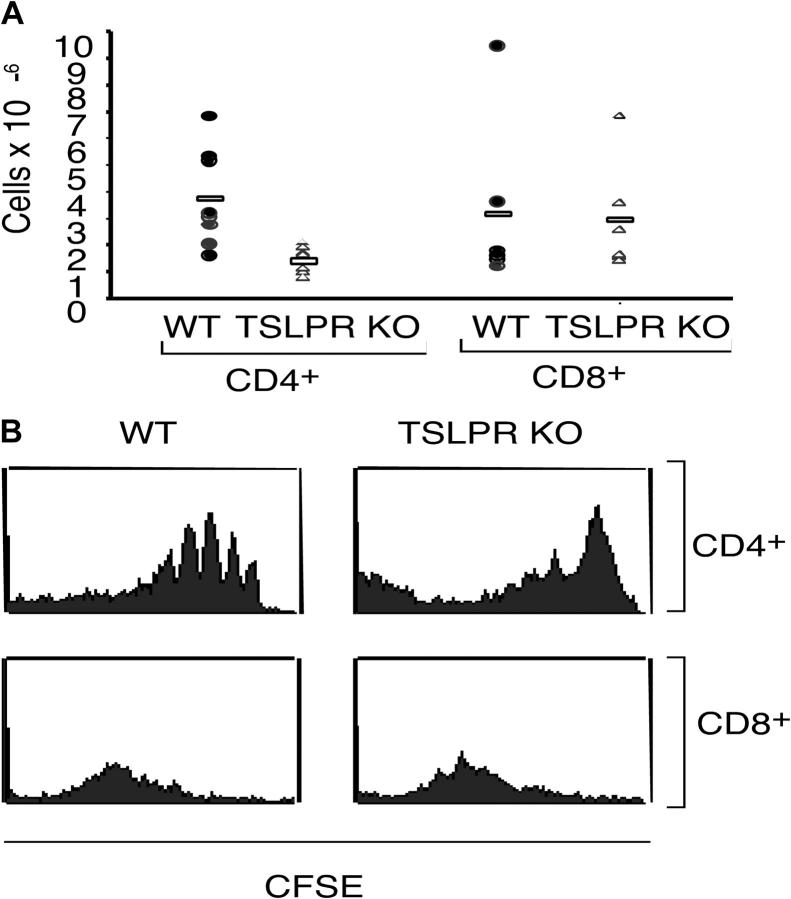
Thymic stromal lymphopoietin (TSLP) signals via a receptor comprising the interleukin (IL)-7 receptor alpha chain and a distinctive subunit, TSLP receptor (TSLPR), which is most related to the common cytokine receptor gamma chain, gamma(c). We have generated TSLPR knockout (KO) mice and found that although these mice had normal lymphocyte numbers, gamma(c)/TSLPR double KO mice had a greater lymphoid defect than gamma(c) KO mice. This indicates that TSLP contributes to lymphoid development and accounts for some of the residual lymphoid development in gamma(c) KO mice and presumably in patients with X-linked severe combined immunodeficiency. Injection of TSLP into gamma(c) KO mice induced the expansion of T and B cells. Moreover, sublethally irradiated TSLPR KO mice showed weaker recovery of lymphocyte populations than wild-type (WT) littermates, even when neutralizing anti-IL-7 antibodies were injected. Interestingly, TSLP preferentially stimulated the proliferation and survival of CD4(+) single positive thymocytes and peripheral T cells in vitro. Additionally, CD4(+) T cells from TSLPR KO mice expanded less efficiently than WT CD4(+) T cells in irradiated hosts, and TSLP preferentially expanded CD4(+) T cells both in vitro and in vivo. Thus, as compared with other known cytokines, TSLP is distinctive in exhibiting a lineage preference for the expansion and survival of CD4(+) T cells.
Figures
References
-
- Leonard, W.J. 2003. Type I Cytokines and Interferons and Their Receptors. Fundamental Immunology. W.E. Paul, editor. Lippincott-Williams and Williams, Philadelphia. 701–747.
-
- Puel, A., S.F. Ziegler, R.H. Buckley, and W.J. Leonard. 1998. Defective IL7R expression in T(−)B(+)NK(+) severe combined immunodeficiency. Nat. Genet. 20:394–397. - PubMed
-
- Friend, S.L., S. Hosier, A. Nelson, D. Foxworthe, D.E. Williams, and A. Farr. 1994. A thymic stromal cell line supports in vitro development of surface IgM+ B cells and produces a novel growth factor affecting B and T lineage cells. Exp. Hematol. 22:321–328. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
