

Mood stabilizer valproate promotes ERK pathway-dependent cortical neuronal growth and neurogenesis
- PMID: 15269271
- PMCID: PMC6729884
- DOI: 10.1523/JNEUROSCI.5747-03.2004
Mood stabilizer valproate promotes ERK pathway-dependent cortical neuronal growth and neurogenesis
Abstract
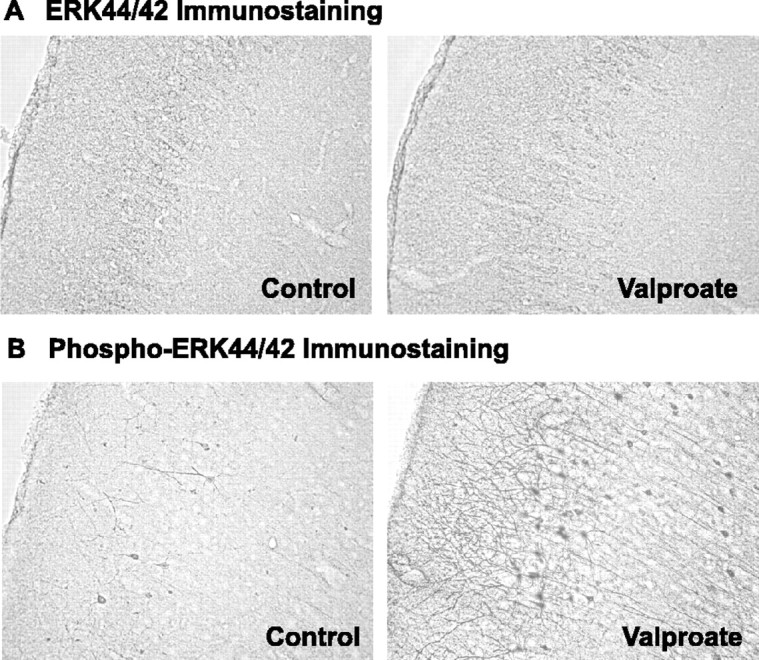
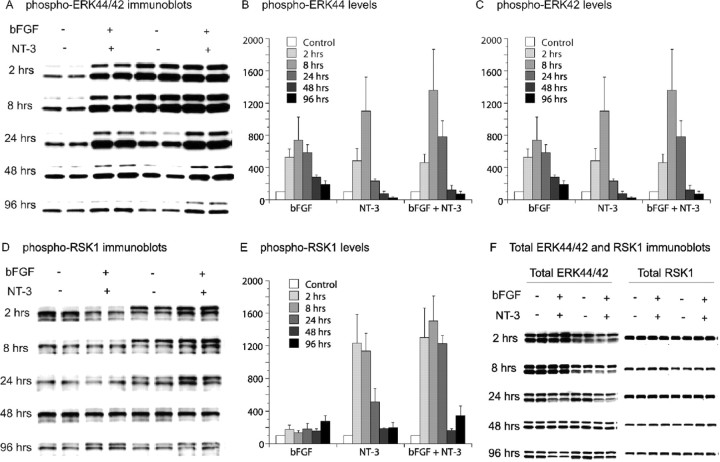
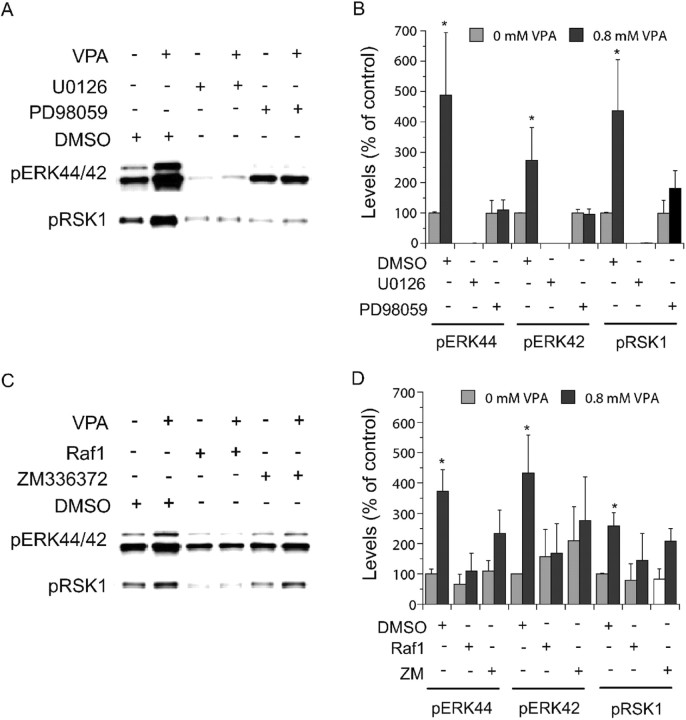
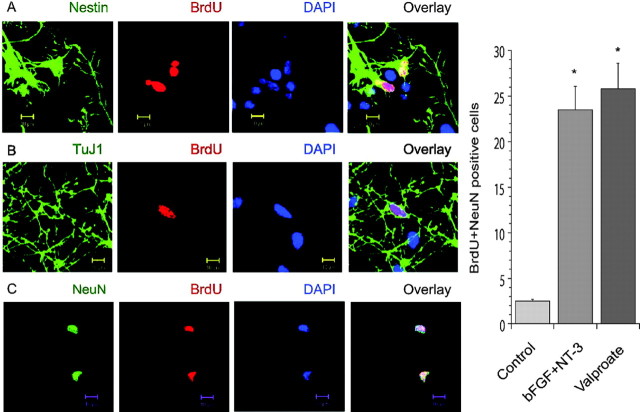
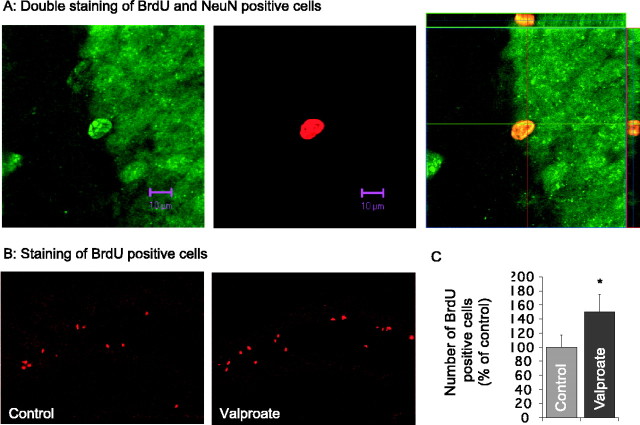
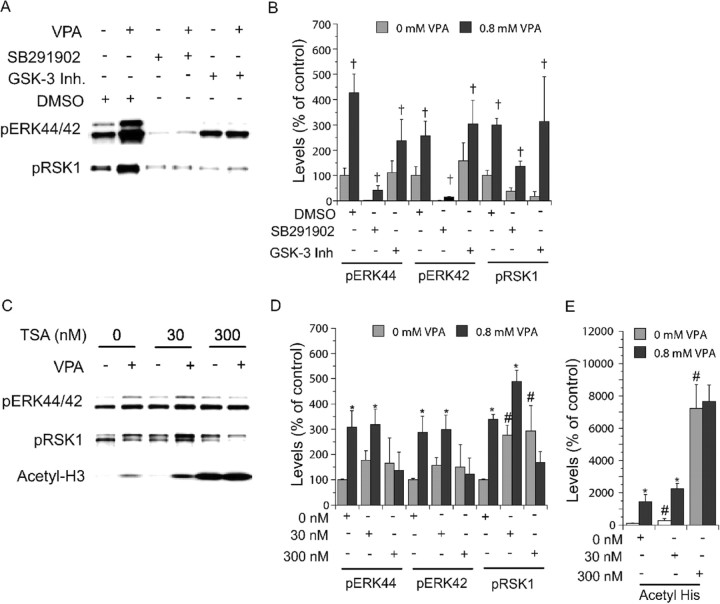
Manic-depressive illness has been conceptualized as a neurochemical illness. However, brain imaging and postmortem studies reveal gray-matter reductions, as well as neuronal and glial atrophy and loss in discrete brain regions of manic-depressive patients. The roles of such cerebral morphological deficits in the neuropathophysiology and therapeutic mechanisms of manic-depressive illness are unknown. Valproate (2-propylpentanoate) is a commonly used mood stabilizer. The ERK (extracellular signal-regulated kinase) pathway is used by neurotrophic factors to regulate neurogenesis, neurite outgrowth, and neuronal survival. We found that chronic treatment of rats with valproate increased levels of activated phospho-ERK44/42 in neurons of the anterior cingulate, a region in which we found valproate-induced increases in expression of an ERK pathway-regulated gene, bcl-2. Valproate time and concentration dependently increased activated phospho-ERK44/42 and phospho-RSK1 (ribosomal S6 kinase 1) levels in cultured cortical cells. These increases were attenuated by Raf and MEK (mitogen-activated protein kinase/ERK kinase) inhibitors. Although valproate affects the functions of GSK-3 (glycogen synthase kinase-3) and histone deacetylase (HDAC), its effects on the ERK pathway were not fully mimicked by selective inhibitors of GSK-3 or HDAC. Similar to neurotrophic factors, valproate enhanced ERK pathway-dependent cortical neuronal growth. Valproate also promoted neural stem cell proliferation-maturation (neurogenesis), demonstrated by bromodeoxyuridine (BrdU) incorporation and double staining of BrdU with nestin, Tuj1, or the neuronal nuclei marker NeuN (neuronal-specific nuclear protein). Chronic treatment with valproate enhanced neurogenesis in the dentate gyrus of the hippocampus. Together, these data demonstrate that valproate activates the ERK pathway and induces ERK pathway-mediated neurotrophic actions. This cascade of events provides a potential mechanism whereby mood stabilizers alleviate cerebral morphometric deficits associated with manic-depressive illness.
Figures


References
-
- Abdel-Latif AA (2001) Cross talk between cyclic nucleotides and polyphosphoinositide hydrolysis, protein kinases, and contraction in smooth muscle. Exp Biol Med 226: 153-163. - PubMed
-
- Altar CA, Whitehead RE, Chen R, Wortwein G, Madsen TM (2003) Effects of electroconvulsive seizures and antidepressant drugs on brain-derived neurotrophic factor protein in rat brain. Biol Psychiatry 54: 703-709. - PubMed
-
- Andresen BT, Rizzo MA, Shome K, Romero G (2002) The role of phosphatidic acid in the regulation of the Ras/MEK/Erk signaling cascade. FEBS Lett 531: 65-68. - PubMed
-
- Apfel SC (2002) Is the therapeutic application of neurotrophic factors dead? Ann Neurol 51: 8-11. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous