

Gamma interferon production, but not perforin-mediated cytolytic activity, of T cells is required for prevention of toxoplasmic encephalitis in BALB/c mice genetically resistant to the disease
- PMID: 15271900
- PMCID: PMC470633
- DOI: 10.1128/IAI.72.8.4432-4438.2004
Gamma interferon production, but not perforin-mediated cytolytic activity, of T cells is required for prevention of toxoplasmic encephalitis in BALB/c mice genetically resistant to the disease
Abstract
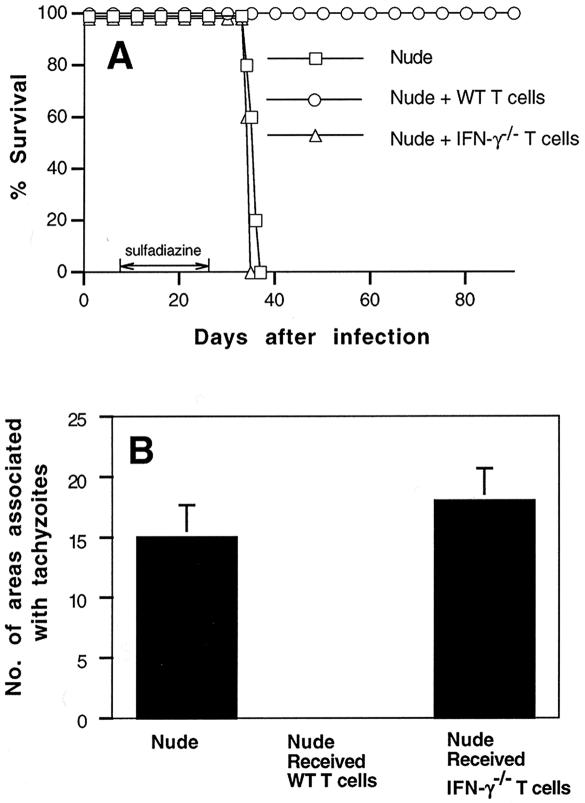
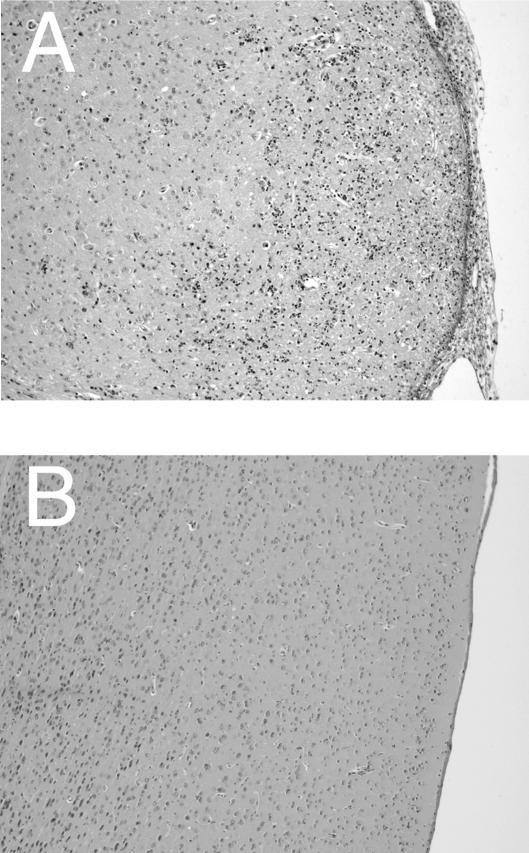
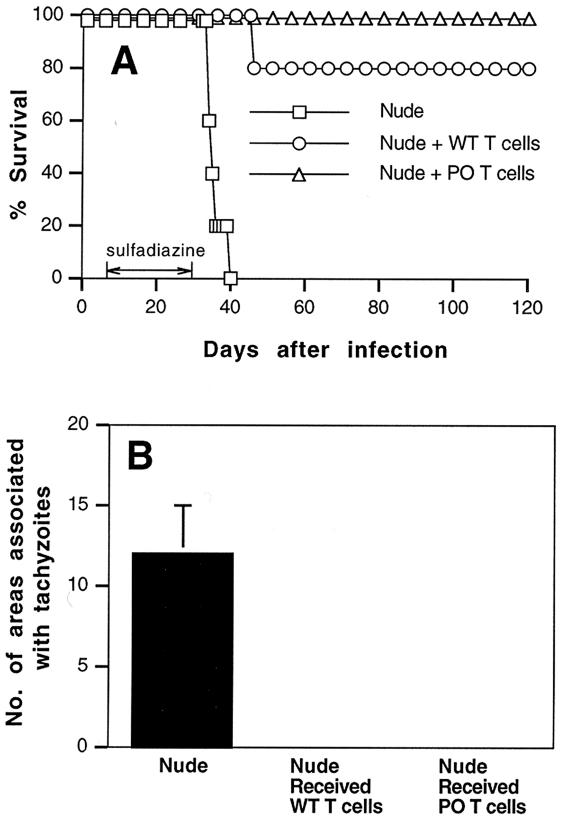
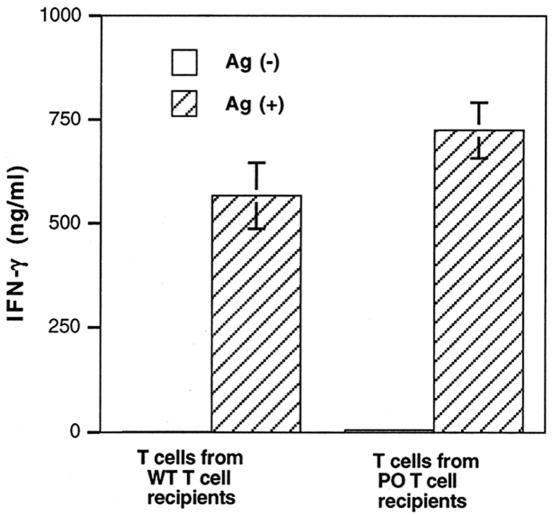
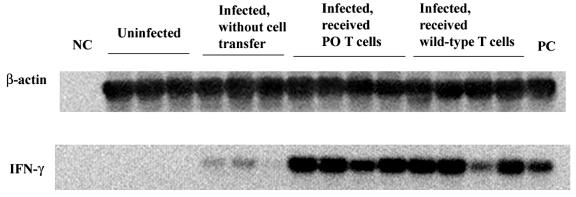
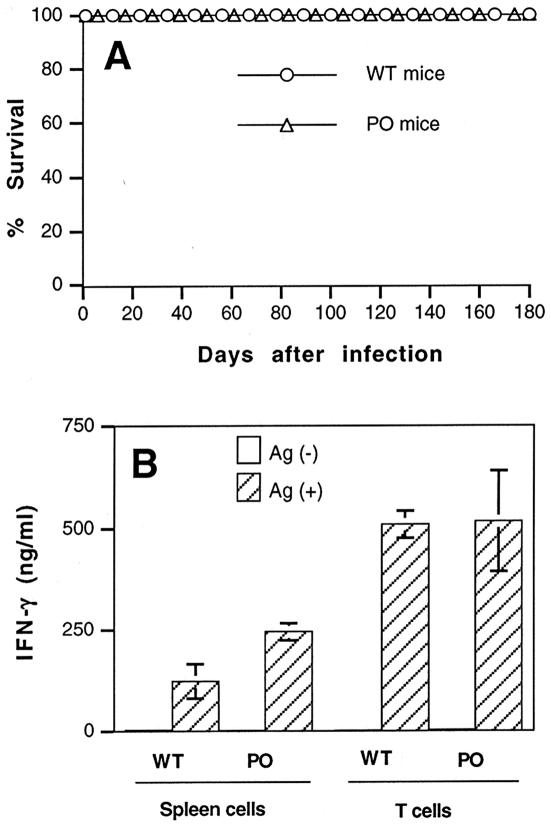
We previously showed the requirement of both T cells and gamma interferon (IFN-gamma)-producing non-T cells for the genetic resistance of BALB/c mice to the development of toxoplasmic encephalitis (TE). In order to define the role of IFN-gamma production and the perforin-mediated cytotoxicity of T cells in this resistance, we obtained immune T cells from spleens of infected IFN-gamma knockout (IFN-gamma(-/-)), perforin knockout (PO), and wild-type BALB/c mice and transferred them into infected and sulfadiazine-treated athymic nude mice, which lack T cells but have IFN-gamma-producing non-T cells. Control nude mice that had not received any T cells developed severe TE and died after discontinuation of sulfadiazine treatment due to the reactivation of infection. Animals that had received immune T cells from either wild-type or PO mice did not develop TE and survived. In contrast, nude mice that had received immune T cells from IFN-gamma(-/-) mice developed severe TE and died as early as control nude mice. T cells obtained from the spleens of animals that had received either PO or wild-type T cells produced large amounts of IFN-gamma after stimulation with Toxoplasma gondii antigens in vitro. In addition, the amounts of IFN-gamma mRNA expressed in the brains of PO T-cell recipients did not differ from those in wild-type T-cell recipients. Furthermore, PO mice did not develop TE after infection, and their IFN-gamma production was equivalent to or higher than that of wild-type animals. These results indicate that IFN-gamma production, but not perforin-mediated cytotoxic activity, by T cells is required for the prevention of TE in genetically resistant BALB/c mice.
Figures
Similar articles
-
Importance of CD8(+)Vbeta8(+) T cells in IFN-gamma-mediated prevention of toxoplasmic encephalitis in genetically resistant BALB/c mice.J Interferon Cytokine Res. 2005 Jun;25(6):338-44. doi: 10.1089/jir.2005.25.338. J Interferon Cytokine Res. 2005. PMID: 15957957
-
TCR V beta 8+ T cells prevent development of toxoplasmic encephalitis in BALB/c mice genetically resistant to the disease.J Immunol. 2003 Apr 15;170(8):4254-9. doi: 10.4049/jimmunol.170.8.4254. J Immunol. 2003. PMID: 12682259
-
Requirement of non-T cells that produce gamma interferon for prevention of reactivation of Toxoplasma gondii infection in the brain.Infect Immun. 2001 May;69(5):2920-7. doi: 10.1128/IAI.69.5.2920-2927.2001. Infect Immun. 2001. PMID: 11292707 Free PMC article.
-
Immunopathogenesis of cerebral toxoplasmosis.J Infect Dis. 2002 Dec 1;186 Suppl 2:S234-40. doi: 10.1086/344276. J Infect Dis. 2002. PMID: 12424703 Review.
-
Interferon-gamma- and perforin-mediated immune responses for resistance against Toxoplasma gondii in the brain.Expert Rev Mol Med. 2011 Oct 4;13:e31. doi: 10.1017/S1462399411002018. Expert Rev Mol Med. 2011. PMID: 22005272 Free PMC article. Review.
Cited by
-
Predominant interferon-γ-mediated expression of CXCL9, CXCL10, and CCL5 proteins in the brain during chronic infection with Toxoplasma gondii in BALB/c mice resistant to development of toxoplasmic encephalitis.J Interferon Cytokine Res. 2010 Sep;30(9):653-60. doi: 10.1089/jir.2009.0119. J Interferon Cytokine Res. 2010. PMID: 20626297 Free PMC article.
-
Immunization with the amino-terminus region of dense granule protein 6 (GRA6) of Toxoplasma gondii activates CD8+ cytotoxic T cells capable of removing tissue cysts of the parasite through antigen presentation by human HLA-A2.1.Microbes Infect. 2023 Nov-Dec;25(8):105182. doi: 10.1016/j.micinf.2023.105182. Epub 2023 Jul 8. Microbes Infect. 2023. PMID: 37423326 Free PMC article.
-
IFNs in host defence and parasite immune evasion during Toxoplasma gondii infections.Front Immunol. 2024 Feb 7;15:1356216. doi: 10.3389/fimmu.2024.1356216. eCollection 2024. Front Immunol. 2024. PMID: 38384452 Free PMC article. Review.
-
MyD88 is crucial for the development of a protective CNS immune response to Toxoplasma gondii infection.J Neuroinflammation. 2013 Feb 1;10:19. doi: 10.1186/1742-2094-10-19. J Neuroinflammation. 2013. PMID: 23374751 Free PMC article.
-
Location of the CD8 T cell epitope within the antigenic precursor determines immunogenicity and protection against the Toxoplasma gondii parasite.PLoS Pathog. 2013;9(6):e1003449. doi: 10.1371/journal.ppat.1003449. Epub 2013 Jun 20. PLoS Pathog. 2013. PMID: 23818852 Free PMC article.
References
-
- Binder, G. K., and D. E. Griffin. 2001. Interferon-γ-mediated site-specific clearance of alphavirus from CNS neurons. Science 293:303-306. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
