

Disruption of the Aspergillus fumigatus gene encoding nucleolar protein CgrA impairs thermotolerant growth and reduces virulence
- PMID: 15271935
- PMCID: PMC470587
- DOI: 10.1128/IAI.72.8.4731-4740.2004
Disruption of the Aspergillus fumigatus gene encoding nucleolar protein CgrA impairs thermotolerant growth and reduces virulence
Abstract
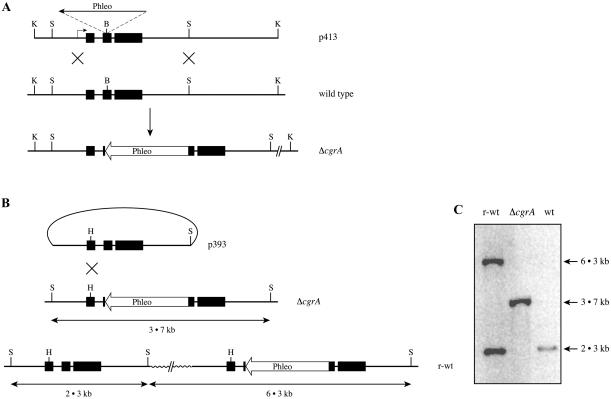
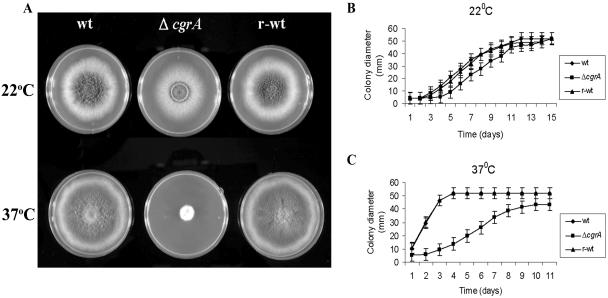
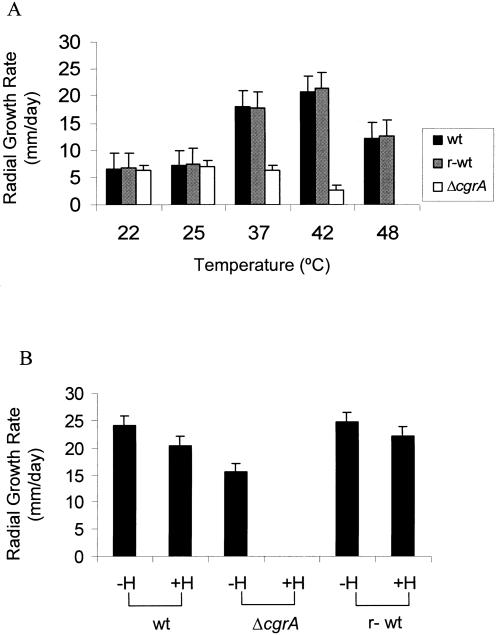
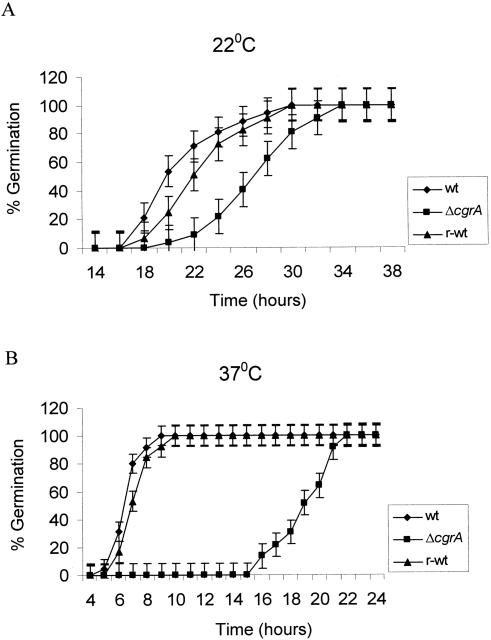
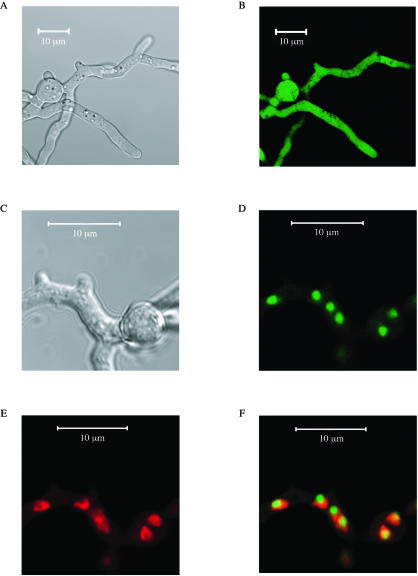
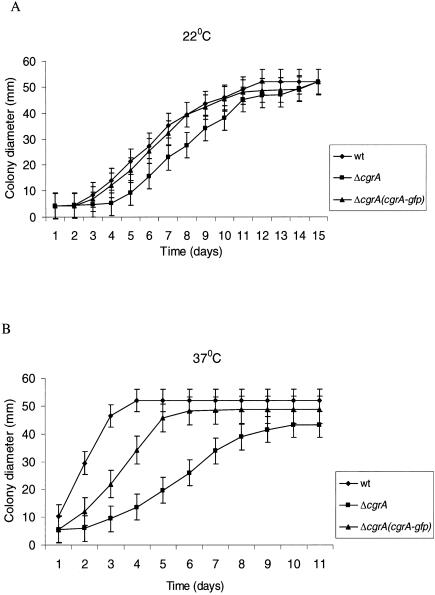
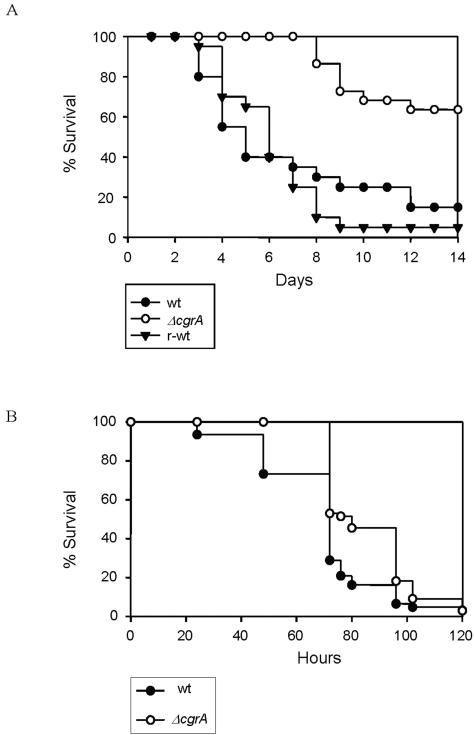
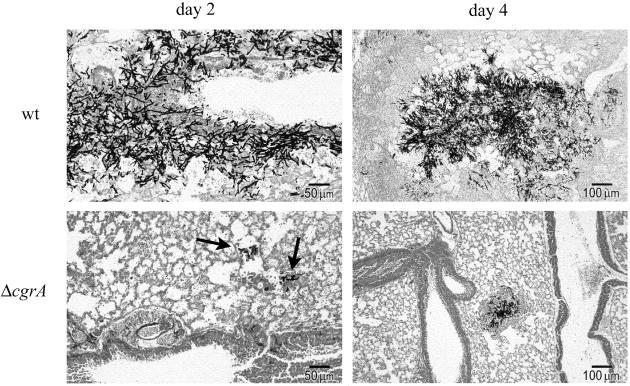
Aspergillus fumigatus CgrA is the ortholog of a yeast nucleolar protein that functions in ribosome synthesis. To determine how CgrA contributes to the virulence of A. fumigatus, a Delta cgrA mutant was constructed by targeted gene disruption, and the mutant was reconstituted to wild type by homologous introduction of a functional cgrA gene. The Delta cgrA mutant had the same growth rate as the wild type at room temperature. However, when the cultures were incubated at 37 degrees C, a condition that increased the growth rate of the wild-type and reconstituted strains approximately threefold, the Delta cgrA mutant was unable to increase its growth rate. The absence of cgrA function caused a delay in both the onset and rate of germination at 37 degrees C but had little effect on germination at room temperature. The Delta cgrA mutant was significantly less virulent than the wild-type or reconstituted strain in immunosuppressed mice and was associated with smaller fungal colonies in lung tissue. However, this difference was less pronounced in a Drosophila infection model at 25 degrees C, which correlated with the comparable growth rates of the two strains at this temperature. To determine the intracellular localization of CgrA, the protein was tagged at the C terminus with green fluorescent protein, and costaining with propidium iodide revealed a predominantly nucleolar localization of the fusion protein in living hyphae. Together, these findings establish the intracellular localization of CgrA in A. fumigatus and demonstrate that cgrA is required for thermotolerant growth and wild-type virulence of the organism.
Figures
References
-
- Alspaugh, J. A., L. M. Cavallo, J. R. Perfect, and J. Heitman. 2000. RAS1 regulates filamentation, mating and growth at high temperature of Cryptococcus neoformans. Mol. Microbiol. 36:352-365. - PubMed
-
- Askew, D. S., C. Bartholomew, A. M. Buchberg, M. B. Valentine, N. A. Jenkins, N. G. Copeland, and J. N. Ihle. 1991. His-1 and His-2: identification and chromosomal mapping of two commonly rearranged sites of viral integration in a myeloid leukemia. Oncogene 6:2041-2047. - PubMed
-
- Ballance, D. J. 1986. Sequences important for gene expression in filamentous fungi. Yeast 2:229-236. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
