

Ca(2+) and K(+) (BK) channels in chick hair cells are clustered and colocalized with apical-basal and tonotopic gradients
- PMID: 15272029
- PMCID: PMC1665200
- DOI: 10.1113/jphysiol.2004.069856
Ca(2+) and K(+) (BK) channels in chick hair cells are clustered and colocalized with apical-basal and tonotopic gradients
Abstract
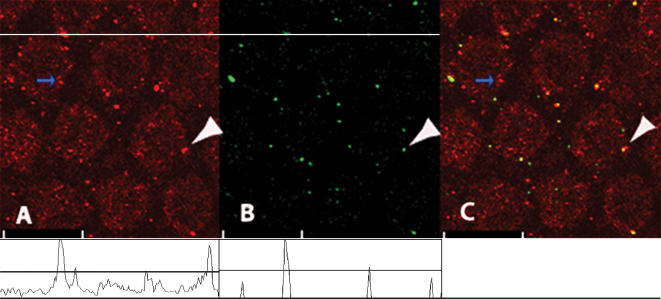
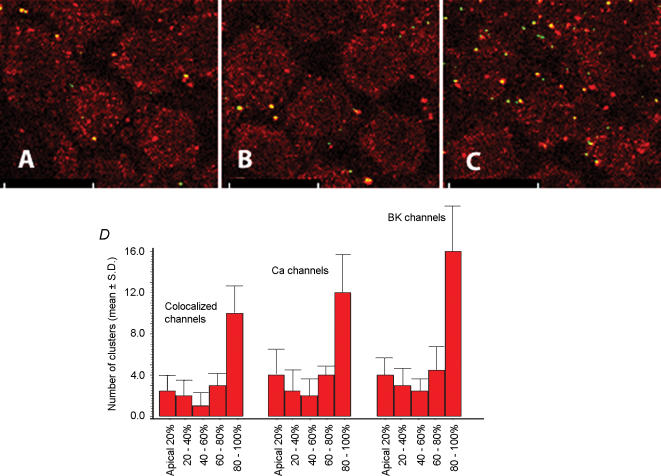
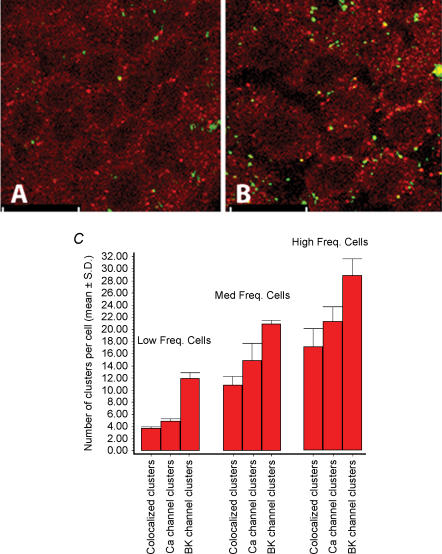
Electrical resonance is a mechanism used by birds and many vertebrates to discriminate between frequencies of sound, and occurs when the intrinsic oscillation in the membrane potential of a specific hair cell corresponds to a specific stimulus sound frequency. This intrinsic oscillation results from an interplay between an inward Ca(2+) current and the resultant activation of a hyperpolarizing Ca(2+)-activated K(+) current. These channels are predicted to lie in close proximity owing to the fast oscillation in membrane potential. The interplay of these channels is widespread in the nervous system, where they perform numerous roles including the control of synaptic release, burst frequency and circadian rhythm generation. Here, we used confocal microscopy to show that these two ion channels are clustered and colocalized in the chick hair cell membrane. The majority of Ca(2+) channels were colocalized while the proportion of colocalized BK channels was markedly less. In addition, we report both an apical-basal gradient of these clusters in individual hair cells, as well as a gradient in the number of clusters between hair cells along the tonotopic axis. These results give physical confirmation of previous predictions. Since the proportion of colocalized channels was a constant function of Ca(2+) channels, and not of BK channels, these results suggest that their colocalization is determined by the former. The molecular mechanisms underpinning their clustering and colocalization are likely to be common to other neuronal cells.
Figures
Similar articles
-
Variation in large-conductance, calcium-activated potassium channels from hair cells along the chicken basilar papilla.J Physiol. 2003 Mar 1;547(Pt 2):357-71. doi: 10.1113/jphysiol.2002.029785. Epub 2003 Jan 17. J Physiol. 2003. PMID: 12562934 Free PMC article.
-
Modeling hair cell tuning by expression gradients of potassium channel beta subunits.Biophys J. 2002 Jan;82(1 Pt 1):64-75. doi: 10.1016/S0006-3495(02)75374-5. Biophys J. 2002. PMID: 11751296 Free PMC article.
-
Calcium-induced calcium release in proximity to hair cell BK channels revealed by PKA activation.Physiol Rep. 2020 Aug;8(15):e14449. doi: 10.14814/phy2.14449. Physiol Rep. 2020. PMID: 32748549 Free PMC article.
-
Mechanisms of hair cell tuning.Annu Rev Physiol. 1999;61:809-34. doi: 10.1146/annurev.physiol.61.1.809. Annu Rev Physiol. 1999. PMID: 10099711 Review.
-
Endothelium as target for large-conductance calcium-activated potassium channel openers.Acta Biochim Pol. 2009;56(3):393-404. Epub 2009 Sep 14. Acta Biochim Pol. 2009. PMID: 19753330 Review.
Cited by
-
Functional coupling between large-conductance potassium channels and Cav3.2 voltage-dependent calcium channels participates in prostate cancer cell growth.Biol Open. 2013 Jul 26;2(9):941-51. doi: 10.1242/bio.20135215. eCollection 2013. Biol Open. 2013. PMID: 24143281 Free PMC article.
-
β4-subunit increases Slo responsiveness to physiological Ca2+ concentrations and together with β1 reduces surface expression of Slo in hair cells.Am J Physiol Cell Physiol. 2011 Mar;300(3):C435-46. doi: 10.1152/ajpcell.00449.2010. Epub 2010 Dec 22. Am J Physiol Cell Physiol. 2011. PMID: 21178105 Free PMC article.
-
Distribution of high-conductance calcium-activated potassium channels in rat vestibular epithelia.J Comp Neurol. 2009 Nov 10;517(2):134-45. doi: 10.1002/cne.22148. J Comp Neurol. 2009. PMID: 19731297 Free PMC article.
-
Molecular Choreography and Structure of Ca2+ Release-Activated Ca2+ (CRAC) and KCa2+ Channels and Their Relevance in Disease with Special Focus on Cancer.Membranes (Basel). 2020 Dec 15;10(12):425. doi: 10.3390/membranes10120425. Membranes (Basel). 2020. PMID: 33333945 Free PMC article. Review.
-
Time-Dependent Effects of Ethanol on BK Channel Expression and Trafficking in Hippocampal Neurons.Alcohol Clin Exp Res. 2015 Sep;39(9):1619-31. doi: 10.1111/acer.12808. Epub 2015 Aug 6. Alcohol Clin Exp Res. 2015. PMID: 26247146 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous